2014.2
BIOLOGIA CELULAR
Prof°. João Paulo
1
APRESENTAÇÃO DA DISCIPLINA
CARO (A) EDUCANDO (A).
A possibilidade de estudar a Biologia Celular nos dias atuais é uma oportunidade que temos para
compreender os fenômenos da vida e vivermos mais e melhor.
A tecnologia sendo bem utilizada nos favorece, em nossos mais diversos anseios, no entanto é
importante pensar e saber pensar sobre esse assunto e sua aplicação para nossa vida. O conhecimento é a
base que nos possibilitará melhor qualidade de vida e a Biologia é uma área em especial que tem tudo a ver
com o acontecimento da vida na terra. O estudo da biologia celular nos proporciona o conhecimento de
como a vida acontece, informação importantíssima para o estudante de técnico em enfermagem. É neste
espaço onde o mesmo vai adquirir uma parcela importante dos conhecimentos necessários ao bom
desempenho do seu papel.
Estudaremos então o conceito de CITOLOGIA: As Hipóteses de origem e evolução;
A Célula: Membrana, Citoplasma e Núcleo;
Orgânulos Celulares;
Divisão Celular: conceitos que determinam a continuação da vida.
Desejamos a todos, discernimento, iniciativa e grandes conquistas.
Abraço e bons estudos.
Educador: João Paulo Silva dos Anjos
ORIGEM E EVOLUÇÃO DAS CÉLULAS
As células eucarióticas teriam ocorrido a partir das células procarióticas que sofreram "dobras" na
membrana plasmática. Esses dobramentos teriam dado origem a "compartimentos" dentro do citoplasma,
que teriam originando as várias estruturas citoplasmáticas delimitadas por membranas- as organelas
membranosas, esses dobramentos também teriam originado a membrana nuclear, que recebe o nome
de carioteca. Isso é o que propõe a hipótese de Robertson sobre a origem das células eucarióticas - a teoria
da invaginação da membrana.
Dentre as organelas membranosas, apenas as mitocôndrias e os cloroplastos parecem ter tido origem
diferente. As mitocôndrias são responsáveis pela respiração celular, ocorrendo em praticamente todo
eucarionte. Os cloroplastos são responsáveis pela fotossíntese, estando presente apenas nas plantas e nos
eucariontes fotossintetizante. A origem de mitocôndrias e cloroplastos encontra fundamento na teoria da
endossimbiose. A teoria da endossibiose foi criada por Lynn Margulis, da universidade de Massachusetts Amherst em 1981, e conhecida com a publicação do ensaio symbiosis in cell evolution ("simbiose na
evolução das células"). Esta teoria propõe que organelas que compõem as eucelula tenham surgido como
consequência de uma associação simbiótica estável entre organismo. Mais especificamente, essa teoria
postula que os cloroplastos e as mitocôndrias dos organismos dos eucariontes têm origem num procarionte
autotrófico - provavelmente um antepassado das cianobacterias atuais - que os viveu em simbiose dentro
de outro organismo, também unicelular, mas provavelmente de maiores dimensões, obtendo assim
proteção e fornecendo ao hospedeiro a energia fornecida pela fotossíntese.
A teoria endossimbiótica de Lynn margulis sugere que as células eucarióticas, nasceram como
comunidades de organismo em interação, que numa célula hospedeira, quer por ingestão, ou como
parasita, com o tempo os elementos originais teriam desenvolvido uma simbiose obrigatória. Margulis
sugeriu ainda que as relações simbióticas sejam uma das principais forças no processo evolutivo, tendo
2
afirmado (em margulis e Sagan, 1996)que "os seres vivos não ocuparam o mundo pela força, mas por
cooperação" e considera a teoria de Darwin de ser a competição a principal força na evolução.
Os primeiros passos rumo à descoberta de como ocorreu a origem e posterior evolução das células
até chegar a forma como elas se encontram hoje foi dado por Friedrich Wöhler em 1832, em um dos
primeiros experimentos de Química Pré-biótica; experimentos estes que tem a finalidade de simular
processos que poderiam ter ocorrido sobre a terra primitiva. Em seu trabalho, Wöhler demonstrou que o
aquecimento de um composto inorgânico, o cianeto de amônia, leva à formação de um composto orgânico,
a uréia, como ilustrado na Fig. 1.
Fig. 1: Formação de uréia a partir de cianeto de amônia. Outras reações demonstraram que o
aminoácido glicina pode ser formado a partir do ácido cianídrico e açúcares podem vir a ser formados a
partir de formaldeído, e ambos estes precursores já foram obtidos a partir de compostos inorgânicos, o que
reforça que todos estes processos podem ter vindo a ocorrer nos estágios iniciais da origem da vida.
Também Oparin e Haldane, sugeriram que durante a evolução dos primeiros organismos vivos a
atmosfera terrestre deveria ter sido redutora e que provavelmente eles teriam surgido de uma mistura
de compostos orgânicos formados em tal atmosfera. Realmente, sabe-se que a síntese de compostos
bioquímicos ocorre muito facilmente em uma atmosfera redutora, mas não em uma atmosfera oxidante.
Por exemplo, se o nitrogênio é introduzido numa atmosfera redutora, será parcialmente convertido em
amônia (NH3). Quem conseguiu demonstrar as previsões de Oparin e Haldane foi Stanley Miller em 1953,
quando submeteu uma mistura de metano, amônia, hidrogênio e água à ação de uma descarga elétrica,
conseguindo assim a formação de vários compostos orgânicos, incluindo aminoácidos. O equipamento
utilizado por Miller é ilustrado na figura 2.
“No frasco pequeno ele colocou água e no resto do aparelho uma mistura de metano, hidrogênio e
amônia.
A mistura gasosa e mais algum vapor d’água foram postos a circular, passando através dos
eletrodos de tungstênio, em conseqüência da fervura da água líquida no frasco pequeno. Então, uma
descarga de centelha foi produzida entre os eletrodos, mediante a aplicação de um alto potencial elétrico
através deles. Os produtos formados na descarga elétrica dissolveram-se na água que se liquefazia no
condensador, e foram transportados até o pequeno frasco”1 . A preferência por essa mistura é devido ao
fato de que era provável que a atmosfera primitiva era formada por esses gases; a água fervendo no frasco
pequeno representava o oceano primitivo (embora seja fato que na época da evolução das primeiras
células o oceano não estava fervendo, ele preferiu representá-lo nessa forma, pois esse procedimento
acelera várias reações, maximizando assim o tempo, pois os resultados desse experimento na realidade
levaram várias centenas de anos para ocorrer sobre a terra primitiva) e as descargas elétricas representaria
os relâmpagos.
3
Fig. 2: Equipamento utilizado por Miller em seu experimento
Várias descobertas estimuladas pelo trabalho de Miller foram sendo feitas sobre quais condições
podem ter se formado as primeiras moléculas orgânicas de importância biológica. Por exemplo, descobriuse que a natureza da mistura de gases não importa desde que seja de caráter redutor; que sempre são
formados conjuntos de moléculas orgânicas similares quando esses gases de caráter redutor são
submetidos a descargas elétricas; que além da glicina, o aquecimento do ácido cianídrico e formaldeído em
uma solução aquosa formam diversos outros aminoácidos e o mais surpreendente, forma também
a adenina, uma das quatro bases dos ácidos nucléicos e um dos componentes do ATP; que a citosina, outra
base nitrogenada, pode ser formada a partir do cianoacetileno, um gás muito similar ao ácido cianídrico e
que a desoxirribose e ribose, os açúcares constituintes dos nucleotídeos, são formados em condições prébióticas quando o formaldeído é agitado com giz ou cal. Assim, pode-se concluir que os constituintes
básicos para a formação do aparato genético estava presente sobre a terra primitiva, e sendo estes os
mesmos compostos básicos na bioquímica moderna, devemos acreditar que as primeiras células surgiram
dessa sopa pré-biótica e que todos os organismos modernos evoluíram a partir dessa primeira célula sem
muitas mudanças em sua composição.
A próxima etapa para a origem das primeiras células deve ter sido a capacidade desses compostos
orgânicos que formavam a sopa pré-biótica, se combinarem para formarem polímeros. Sabe-se através
de experimentos que não é possível que se formem polipeptídios e polinucleotídios com facilidade através
de soluções diluídas de aminoácidos e nucleotídios. Por isso, acredita-se que a formação de polímeros deve
ter ocorrido em uma sopa pré-biótica concentrada. Os principais mecanismos de concentração que
atuaram na terra primitiva favorecendo a formação dos polímeros foram a evaporação, o congelamento, a
adsorção e a formação de gotículas coloidais ou coacervados. Todas essas formas são plausíveis de terem
ocorrido sobre a terra primitiva, porém, ainda não se pode afirmar com certeza qual teria sido a mais
importante. A evaporação é um mecanismo muito comum que ocasiona a concentração de soluções
aquosas diluídas, podendo até formar depósitos sólidos se permanecerem por muito tempo. Assim, parece
provável que a evaporação tenha sido um importante mecanismo que concentrava soluções em poças
deixadas pelas marés, por exemplo. Alternativamente, o congelamento também deve ter atuado,
principalmente por ser possível através de ele concentrar materiais voláteis. Mas sem dúvidas, os dois
mecanismos principais que ocasionaram a concentração das moléculas para o estágio final da origem
celular (a evolução da membrana que será tratado mais adiante), foi a adsorção e formação de
coacervados. Sabe-se que moléculas pré-bióticas importantes aderem (adsorvem) à superfície de minerais
e que fosfatos minerais catalisam reações pré-bióticas. Portanto, é muito provável que os minerais
4
extraiam substâncias da sopa pré-biótica diluída e as concentrava em sua superfície onde poderia também
ter ocorrido importante reações através de catalisadores minerais. A formação de partículas coloidais
(conservados) ocorre quando compostos como sabões e detergentes no contato com a água não formam
soluções verdadeiras, eles se dispersam e formam gotículas de matéria orgânica. Oparin propôs que as
gotículas de conservados devem ter sido o melhor local para as reações de polimerização, pois muitas
reações de condensação (polimerização) ocorrem mais facilmente em meios não aquosos. Porém, é difícil
compreender por que as condensações a seco foram substituídas pelas aquosas no decorrer da evolução
celular. O fato é que as condensações térmicas quando ocorrem, formam boas quantidades de peptídeos,
pois quando uma solução de aminoácidos é fortemente aquecida, a água é eliminada das ligações e então
se formam os polímeros. Essa seria uma explicação ideal para a formação dos polímeros, pois não é
necessária a participação de reagentes, se não fosse o fato de que a reação não ocorre em temperaturas
inferiores a 130ºC. Como é pouco provável que tenha ocorrido temperaturas superiores aos 80ºC na terra
primitiva, então seria necessária a ajuda de um catalisador orgânico ou mineral simples que permitisse a
polimerização nas temperaturas entre 70 e 80ºC. Um catalisador desse tipo poderia ter sido, por exemplo,
a uréia, pois com o auxílio desta é possível adicionar fosfatos aos nucleotídeos, para que polipeptídios
curtos se formem a partir deles.
Alternativamente, a dificuldade encontrada para se obter polímeros formados em soluções aquosas
é devido ao fato de todas as proteínas e ácidos nucléicos serem decompostos em seus elementos
constituintes ao reagirem com a água. E a reação oposta não ocorre, ou seja, aminoácidos e nucleotídeos
não se combinam para formarem proteínas e ácidos nucléicos, respectivamente, a não ser que energia seja
despendida para isso, já que a solução se encontra em equilíbrio químico. Então, deve ter havido
intermediários de alta energia que fornecessem energia ao sistema para que houvesse a reação de
condensação. Porém, como esses intermediários se formaram na terra primitiva e quem eram eles? Uma
resposta para essa pergunta é que os fosfatos de alta-energia já estavam presentes desde o início da
evolução do aparato genético, sendo formados a partir de nucleotídeos e fosfatos inorgânicos na presença
de uréia ou outro catalisador. Por isso, há pesquisadores que acreditam que ambos os tipos de
condensação – térmica (a seco) e em solução (aquosa) - ocorreram sobre a terra primitiva e que somente
as em solução evoluíram para estarem presentes hoje nas condensações biológicas modernas. Outros
acreditam que talvez somente as condensações em solução ocorressem e que ainda não foram descobertos
os mecanismos eficientes a partir dos quais se condensem os monômeros em polímeros nessas soluções
aquosas. O fato é que somente as condensações em solução existem nos sistemas biológicos modernos.
O próximo passo crucial que marcou decisivamente a passagem de um sistema onde ocorriam reações
para a formação de moléculas orgânicas poliméricas simples para um sistema biológico auto reprodutor, foi
a seleção de determinados polímeros capazes de dirigir sua própria replicação. Das duas classes principais
de macromoléculas atuais, apenas os ácidos nucléicos são capazes de auto-replicação. Por isso, acredita-se
que os polinucleotídeos foram as primeiras moléculas genéticas sobre a terra primitiva e que os
polipeptídios poderiam ter sido catalisadores importantes, uma vez que é pouco provável que uma
molécula aparentada às proteínas modernas possa ter sido capaz de se auto replicar. Evidências indicam
que os ácidos nucléicos poderiam ter se replicado na ausência de enzimas, uma vez que experimentos
demonstraram a replicação resíduo por resíduo em condições pré-bióticas de polinucleotídios artificiais
que se assemelham aos ácidos nucléicos. Por exemplo, quando o poli U (um polímero que contém apenas
ácido uridílico) é misturado com derivados monoméricos de ácido adenílico (A), a temperaturas
suficientemente baixas, eles têm a capacidade de dirigir a organização de A de maneira a fazê-los formar
uma hélice. Essa conformação em hélice torna a ligação dos resíduos de A muito mais fácil do que quando
os mesmos se encontram livres. Da mesma maneira, já foi observado que o poli C (um polímero que
contém apenas derivados do ácido citidílico) dirige da mesma forma a polimerização de derivados de G. E
5
que C não influi na polimerização de A e que U não influi na polimerização de G. Dessa forma, podem-se ser
visualizadas as mesmas regras propostas por Watson e Crick para o emparelhamento de bases que tornam
a replicação do DNA possível.
A adaptação posterior que levaria à evolução de um sistema mais sofisticado de replicação, do código
genético e do aparato sintetizador de proteínas deve ter envolvido interações entre os primeiros polímeros
replicadores (sejam eles os RNAs, DNAs ou outro tipo de ácido nucléico que teria se adaptado à estrutura
replicadora como já foi proposto) e outras moléculas não replicadoras que se encontravam no seu meio.
“Primeiro, os polinucleotídeos “aprenderam” a capturar qualquer molécula pequena na sopa pré-biótica
que pudesse ajudá-lo a levar a cabo a sua replicação. Mais tarde, devem ter aprendido a unir pequenas
moléculas da sopa pré-biótica e a incorporá-las aos seus produtos poliméricos. A mais importante interação
entre os ácidos nucléicos replicadores e as moléculas menores envolviam os aminoácidos; o código
genético é o produto final da evolução da interação entre o ácido nucléico e o aminoácido. E a invenção da
síntese protéica permitiu aos ácidos nucléicos dirigir a síntese de enzimas e, desta maneira, estabelecer
quase que um controle total sobre o seu meio químico”1 .
Sempre se indaga qual poderia ter sido a molécula replicável ancestral, RNA ou DNA? Evidências
apontam para o RNA como sendo esta primeira molécula capaz de armazenar as informações genéticas,
uma vez que é composto por uma fita simples, enquanto o DNA é uma dupla-fita, e isto leva à necessidade
de enzimas capazes de abrir a dupla-fita para a replicação no DNA enquanto o RNA pode ser lido ou
replicado diretamente; além disso, o RNA pode atuar na catálise de reações, inclusive a polimerização de
nucleotídeos e aminoácidos, o que foi crucial tanto para as reações de replicação das primeiras moléculas
de RNA quanto para a evolução do mecanismo sintetizador de proteínas. As imperfeições nas primeiras
replicações provavelmente fizeram surgir inúmeras variantes de RNAs catalíticos que ao se replicarem em
taxas diferentes, competiam e apenas os que se replicavam mais rápido se saíram vencedores. No início, a
evolução do mecanismo sintetizador de proteínas não deve ter envolvido mais que uns poucos polímeros,
parcialmente ordenados que não devessem ter quase toda a especificidade hoje presente no mecanismo
moderno. Esses poucos polímeros deveriam ter sido: uma molécula de informação muito parecida com o
RNA mensageiro (RNAm), cuja seqüência de nucleotídeos era lida para criar uma seqüência específica de
aminoácidos; houve um grupo de moléculas muito parecidas com o RNA de transferência (RNAt) que
poderia ativar aminoácidos específicos e também deve ter havido um catalisador, muito parecido com o
RNA ribossomal (RNAr) que teria facilitado a junção dos aminoácidos trazidos pelo RNA de transferência a
partir da leitura do RNA de informação. A falta de especificidade desse mecanismo inicial levava a
aminoácidos muito parecidos não terem sido discriminados. Também se acredita que não havia sinais
eficientes de inicio e parada no código genético primitivo. Porém, um aspecto da síntese protéica que foi
muito bem definido desde o início deve ter sido o código de três letras, pois mesmo que se tivesse evoluído
um código com número de letras diferente, isso teria levado a uma interpretação desastrosa da informação
quando houvesse a transição para um código de três letras. A estrutura do código provavelmente é
resultado de um acidente, ou não. Por exemplo, “talvez a glicina interaja mais intensamente com a
seqüência GGG (ou com a seqüência complementar CCC) do que com qualquer outro tri nucleotídeo. Neste
caso GGG codificaria a glicina, se o mecanismo genético viesse a evoluir uma segunda vez.
Alternativamente, a relação entre tri nucleotídeos e aminoácidos, especificada pelo código, poderia ter sido
determinada por fatores arbitrários. A glicina talvez se tivesse concentrado com GGG em alguma poça de
maré em particular, e isto teria levado à sua associação no código . Se foi assim, o código genético,
evoluindo uma segunda vez, possivelmente não associaria GGG à glicina”. Essa fase da evolução da vida em
que o material genético era o RNA é conhecida pelo termo “mundo de RNA”. Um motivo para transição
entre o uso do RNA para o DNA como material genético é devido à limitação imposta pela alta taxa de
mutação da molécula de RNA. Desse modo, a evolução do DNA teria reduzido a taxa de mutação, o que é
6
de extrema importância para uma molécula que armazena as informações genéticas que são passadas
hereditariamente.
Com a evolução da cooperação entre as moléculas, foi de extrema importância a origem de uma
membrana que as envolvesse, uma vez que vantagens que poderiam ser adquiridas com a origem de novas
sínteses de compostos bioquímicos seriam perdidas se fossem compartilhadas com outros grupos de
moléculas competidoras. Neste caso a membrana deveria ser semipermeável, ou seja, impermeável às
pequenas moléculas úteis produzidas no interior da célula, mas permeável às moléculas vindas de fora. Os
mecanismos mais importantes pelos quais as famílias de polímeros foram mantidas juntas em presença de
uma grande quantidade de água foram, como já comentados anteriormente, a adsorção ou a formação de
partículas coloidais. As duas teorias são plausíveis, embora ainda não se possa confirmar com certeza qual
realmente ocorreu, ou se ambas ocorreram, qual teria sido a mais importante. Mas agora, o que realmente
importa é que de alguma forma as moléculas se mantinham unidas e necessitavam de uma delimitação. Se
acreditarmos que fosfolipídios estavam presentes na sopa pré-biótica, não é muito difícil imaginar como
teria se formado a membrana, uma vez que estes possuem a propriedade de formar espontaneamente
bicamadas (Fig. 3) pela aproximação de suas caudas hidrófobas, deixando suas cabeças hidrófilas em
contato com a água. Esse arranjo poderia ter envolvido esses conjuntos de moléculas que formavam
agregados, separando o seu interior do ambiente externo pela formação da bicamada lipídica. Assim, teria
se formado as células, a partir de conjuntos cooperativos de polímeros envolvidos por uma bicamada de
lipídeos
.
Fig. 3: Conformação espontânea adquirida pelos fosfolipídios em soluções aquosas
No início do desenvolvimento das primeiras células, todos os compostos bioquímicos essenciais
estavam disponíveis na sopa pré-biótica para serem consumidos. Porém, esses recursos foram se
esgotando com o passar do tempo, e somente o desenvolvimento da síntese dos compostos pelas células
seria capaz de suprir as demandas energéticas. As vias bioquímicas que foram desenvolvidas pelas células
diferiam pouco das vias pré-bióticas que formavam os compostos bioquímicos de que as células
necessitavam, por exemplo, as vias bioquímicas necessitam da participação de enzimas. “Acredita-se que
muitas vias biossintéticas desenvolveram-se fazendo, paradoxalmente, o caminho inverso da
decomposição espontânea”1 . “A geração e utilização controlada de energia metabólica são fundamentais
para todas as atividades celulares, e as principais vias do metabolismo energético são altamente
conservadas em atuais células”2 , o que reforça a teoria de que todos os organismos descendem de um
ancestral comum. Na atmosfera inicialmente anaeróbica, o primeiro mecanismo de geração de energia
envolvia provavelmente uma via muito parecida com a glicólise atual, que converte glicose em ácido lático
na ausência de oxigênio com ganho líquido de duas moléculas de ATP. A evolução do mecanismo
fotossintético (Fig. 4) foi o próximo passo na evolução do metabolismo celular. Inicialmente, as primeiras
células utilizavam H2S para converter CO2 em moléculas orgânicas, uma via que ainda é utilizada por
algumas bactérias hoje em dia. A utilização de H2O como doador de elétrons para converter CO2 em
7
moléculas orgânicas só surgiu posteriormente, e o subproduto desta reação (oxigênio) foi responsável por
com o passar do tempo se acumular na atmosfera e mudar os rumos da evolução metabólica. Essa
mudança causada no ambiente em que as células estavam evoluindo foi provavelmente a causa do
desenvolvimento do metabolismo oxidativo, uma via muito mais vantajosa pelo ganho líquido maior de
moléculas de ATP por molécula de glicose.
Fig. 4: Descrição simplificada da fotossíntese
“A divisão classificatória mais inicial da vida celular é uma árvore trifurcada em arques, bactérias e
eucariotos. As arques e bactérias são procariotos, e ambas existiam na terra há 2 e 3 bilhões de anos. O
outro tipo de célula, a eucariótica, evoluiu depois dos procariotos”3 . A datação mais antiga da origem das
células eucarióticas provém de fósseis químicos de certas gorduras provavelmente relacionadas ao
metabolismo eucarioto, datados de 2,7 bilhões de anos, porém, como as gorduras podem não ser boas
indicadoras, é mais sensato que a origem dos eucariotos tenha ocorrido entre 2,2 e 1,8 bilhões de anos
como é sugerido pelo relógio molecular. As células procarióticas atuais são versões mais complexas das
primeiras células que surgiram sobre a terra.
A diferença mais conspícua entre células procariotas e eucariotas é a presença de núcleo nas
eucarióticas e ausência dele nas procarióticas. Além disso, células eucariotas possuem organelas, um
processo de divisão celular chamado mitose, um processo ainda mais sofisticado de divisão celular que dá
origem aos gametas, além de muitas outras diferenças. Foi proposto que as células eucarióticas evoluíram à
partir das procarióticas quando estas sofreram invaginações da membrana citoplasmática, dando origem
aos compartimentos internos que formaram as organelas e, inclusive a carioteca, a membrana que delimita
o núcleo. Essa teoria é proposta por Robertson como sendo "a Teoria da Invaginação da Membrana".
Somente as mitocôndrias e os cloroplastos parecem não ter se originado dessa forma. É muito provável
que essas organelas se originaram por simbiose de uma célula procarionte unicelular autotrófica que
possivelmente é um antepassado das cianobactérias atuais com um organismo também unicelular, porém
possivelmente de maiores dimensões. Desse modo a primeira fornecia energia oriunda da fotossíntese e a
segunda proteção. Essa teoria da simbiose foi proposta por Lynn Margulis e é conhecida como "teoria da
endossimbiose". Agora, o próximo passo da evolução será começar a unir as células em cooperação, assim
como uniu os polímeros no início da evolução rumo às primeiras células, para que surjam os primeiros
organismos multicelulares.
8
TEORIA CELULAR
A Teoria Celular, criada por Robert Hooke, em 1665, estabelece a célula como a unidade morfofisiológica
dos seres vivos, ou seja, a célula é a unidade básica da vida. No final da década de 1830,
dois cientistas alemães, Matthias Jakob Schleiden, ex-advogado que abandonou a profissão para estudar a
estrutura e fisiologia das plantas, determinando também que todas as plantas apresentavam organização
celular, e Theodor Schwann, médico dedicado ao estudo da anatomia dos animais, estendeu a teoria de
Mathias aos animais, formulando a hipótese de que todos os seres vivos são constituídos por células,
construindo a base da teoria celular.
IMPORTÂNCIA.
A Teoria Celular foi uma das mais importantes generalizações da história da Biologia. Ficou claro que,
apesar das diferenças quanto à forma e função, todos os seres vivos têm em comum o fato de serem
formados por células. Portanto, para a plena compreensão do fenômeno da vida, é preciso conhecer as
células.
O entusiasmo pela Teoria Celular também levou os biólogos à investigação sobre a origem das células
vivas. Alguns acreditavam que as células se formavam espontaneamente, a partir da aglomeração de
determinados tipos de substâncias, enquanto outros se opunham a essa idéia, afirmando que uma célula
somente podia originar-se de outra célula pré-existente.
A teoria celular foi "o alicerce" para todo o desenvolvimento do estudo biológico. vejam:
Todo ser vivo é formado por células.
9
A vida depende da Autonomia e da integridade da célula.
Toda célula é portadora de material genético, o DNA e o RNA.
A célula é responsável por todo o metabolismo do organismo, em conjunto com outras, forma os
órgãos e sistemas.
Toda célula se origina de outra célula pré-existente. Sendo que elas fazem a reprodução
assexuada.
Nem todos os seres vivos possuem organização celular: os vírus não possuem tal organização, ou seja,
são acelulares e precisam necessariamente invadir células vivas para se reproduzir. Eles são parasitas
intracelulares obrigatórios.
MODELOS DE CÉLULAS DIFERENTES
Célula eucariota: Células eucariotas ou eucariontes ou eucélulas são mais complexas do que as
procarióticas, pois possuem membrana nuclear individualizada e vários tipos de organelas. Uma célula
eucariótica possui verdadeiro núcleo, (núcleo definido e protegido pelo envoltório nuclear) que contém um
ou mais nucléolos. É constituída por muitas organelas citoplasmáticas, ao contrário das células
procarióticas. E podem ser animais ou vegetais. De acordo com os cientistas as células eucarióticas
surgiram a partir das procarióticas, que são células relativamente mais simples.
10
Célula procariota: As células procarióticas ou procariontes são relativamente simples (comparativamente
às eucarióticas) e são as que se encontram nas bactérias e cianófitas ("algas" azuis ou
cianobactérias). Procariotos são organismos unicelulares.
Os procariontes apresentam metabolismos muito diversificados, o que é refletido na sua capacidade de
colonização de diferentes ambientes, tais como tratos digestivos de animais, ambientes vulcânico... Apesar
de não possuirem organelas celulares, podem conduzir seus processos metabólicos na membrana celular.
EXEMPLO DE CÉLULA VEGETAL
11
EXEMPLO DE CÉLULA ANIMAL
Diferenças básicas:
A célula vegetal possui a parede celular e os cloroplastos, organelas ausentes na célula animal:
A célula animal possui os centríolos e os lisossomos, organelas que geralmente não aparecem na célula vegetal.
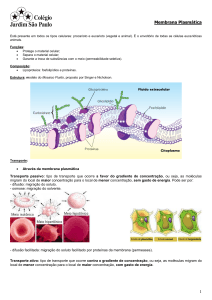
MEMBRANA: Película muito fina e elástica lipoproteica (constituída de proteínas e lipídios) e que envolve
toda a célula. É o envoltório celular.
FUNÇÃO: Controla a passagem de substâncias entre o meio intracelular e extracelular (entrada e saída de
substâncias).
12
OBSERVE:
A MEMBRANA E OS TIPOS DE TRANSPORTE
Processo
Transporte
passivo
Difusão
simples
osmose
Difusão
facilitada
ATRAVÉS
MEMBRANA
DA
Transporte ativo
Condição
Exemplo
Existir
uma
diferença
de
e concentração; a Oxigênio,
célula não gasta carbônico
energia
água
Existir
uma
diferença
de
concentração; a
célula não gasta
energia
Existir
uma
proteína
carregadora e
haver gasto de
energia
por
gás
e
Açúcares
e
aminoácidos
Açúcares,
aminoácidos e
ions de cálcio e
de potássio
13
parte da célula
POR ENVOLVIMENTO
E CAPITURA PELA
MEMBRANA
Endocitose
(ENDOCITOSE
E
EXOCITOSE)
Exocitose
Pinocitose
(partículas
pequenas
e
geralmente
líquidas)
Fagocitose
(partículas
sólidas)
Gotículas
lipídios
de
Vírus, bactérias
e
restos
celulares
Processo em
que
as
substâncias
saem
das
células
FAGOCITOSE:
PINOCITOSE
14
CITOPLASMA: Todo o espaço existente entre a membrana e o núcleo. É onde está presente o
hialoplasma.
HIALOPLASMA: Substância gelatinosa (gel contendo água e proteínas), que facilita a distribuição de
substâncias. É onde estão as organelas citoplasmáticas.
CONHECENDO AS ORGANELAS CITOPLASMÁTICAS OU ORGÂNULOS CELULARES:
15
RETÍCULO ENDOPLASMÁTICO
LISO: Sistema de finas membranas duplas lipoproteicas que forma uma complexa rede espalhada por
toda a célula.
FUNÇÕES:
Transporte de substâncias no interior da célula;
Armazenamento de substâncias produzidas;
Síntese de lipídios;
Participa do metabolismo dos carboidratos;
Participam da neutralização de substancia tóxicas como drogas e venenos.
RETÍCULO ENDOPLASMÁTICO RUGOSO: Sistema de finas membranas duplas lipoproteicas que forma uma
complexa rede espalhada por toda a célula, apresentando ribossomos aderidos em suas membranas.
FUNÇÕES:
Síntese de proteínas devido aos ribossomos aderidos em suas membranas.
SISTEMA GOLGIENSE: Constituído por várias unidades menores (dictiossomos com cinco ou mais sacos
achatados).
16
FUNÇÕES:
Armazenamento de proteínas antes de serem utilizadas pelo organismo;
Secreção celular (muco);
Produção do acrossomo.
LISOSSOMOS: Organela constituída por pequenas vesículas lipoproteicas contendo substâncias necessárias
à digestão celular.
FUNÇÕES: digestão celular.
17
PEROXISSOMOS: Vesículas semelhantes aos lisossomos.
FUNÇÕES: Transforma substâncias tóxicas em substâncias não tóxicas ao organismo.
MITOCÔNDRIA: Organelas membranosas e em forma de bastão. (composta por membrana externa,
membrana interna, cristas mitocondriais e matriz mitocondrial).
FUNÇÕES: Respiração celular (obtenção de energia para a célula).
18
OS PLASTOS: Orgânulos característicos de células vegetais exceto bactérias, cianobactérias e fungos.
Existem os LEUCOPLASTOS = incolor . Ex.: amiloplasto = tecido de reserva dos vegetais .
CROMOPLASTOS=pigmentos que apresenta coloração. Ex.: cloroplastos = pigmento verde.
Obs.: Os cloroplastos são constituídos por:
Membrana externa e interna;
Estroma onde estão os tilacóide e/ou lamela(grana).
19
CENTRÍOLOS: Estruturas cilíndricas formadas por nove túbulos triplos, encontrados tipicamente em células
animais:
FUNÇÕES:
“Orienta” o deslocamento dos cromossomos durante a divisão celular.
Originam cílios e flagelos, permitindo a locomoção da célula ou do organismo no meio líquido. Ex.:
espermatozóides, Paramércio.
CITOESQUELETO: Conjunto de filamentos que formam uma teia tridimensional no interior da célula, tratase de uma rede de microtúbulos.
20
FUNÇÕES: Sustentação e manutenção do formato celular. Ex.: células que não possuem parede celular,
movimento ameboide.
VACÚOLOS: Espaço delimitado por membrana lipoproteica. Digestão celular realizada pelos lisossomos.
21
NÚCLEO CELULAR: É onde estão armazenadas todas as informações sobre a função e a estrutura das
células.
COMPONENTES DO NÚCLEO:
Envoltório nuclear ou carioteca: Membrana com numerosos poros que controla o trânsito de
moléculas entre o núcleo e o citoplasma.
Nucleoplasma: Material gelatinoso que preenche o espaço interno do núcleo.
Nucléolos: Corpúsculos arredondados sem membranas ricos em RNA ribossômico (um tipo de
ácido nucléico), que associados a proteínas servem para formar novos ribossomos.
22
Cromatina: É o material genético que aparece como filamentos finos imersos no nucleoplasma. A
cromatina condensa (enrola), formando os cromossomos, onde estão os genes constituídos por
DNA.
Obs.: Aos vinte e três (23) pares de cromossomos, chamamos de cromossomos homólogos, ou seja, dois
(2) cromossomos de cada tipo.
COMPOSIÇÃO QUÍMICA DA CÉLULA
Substâncias que compõem as células: Todos os seres vivos possuem moléculas e elementos que são
essenciais para a sua composição e para o seu metabolismo. É uma grande variedade de substâncias
orgânicas e inorgânicas que fazem parte dessa composição.
Substâncias orgânicas
Carboidratos ou glicídios (açúcares)
Lipídios (óleos e as gorduras)
Proteínas (aminoácidos)
Vitaminas
Substâncias inorgânicas
Água
Sais minerais
23
SISTEMA ENDOMEMBRANOSO
Detalhe do sistema da endomembrana e seus componentes
Bicapa lipídica: O sistema endomembranoso é composto por membranas diferentes, que estão suspensas
no citoplasma dentro de uma célula eucariótica. Estas membranas dividem a célula em compartimentos
funcionais e estruturais, ou organelas. Em eucariontes, as organelas do sistema endomembranoso incluem:
o envoltório nuclear, retículo endoplasmático, ocomplexo de Golgi, lisossomos, vacúolos, vesículas e
a membrana celular. O sistema é definido com mais precisão como o conjunto de membranas que formam
uma única unidade funcional e de desenvolvimento, quer ser conectados diretamente ou através de troca
de material de transporte vesicular. O sistema endomembranoso não inclui as membranas
das mitocôndrias e cloroplastos.
24
O envoltório nuclear é uma membrana que contém duas camadas, que engloba o conteúdo do núcleo. O
retículo endoplasmático é uma organela sintetizante e transportadora, dentro do citoplasma, em células
animais e vegetais. O aparelho de Golgi é uma série de múltiplos compartimentos, onde moléculas são
"empacotadas para entregar" outros componentes celulares ou pela secreção celular. Em procariontes, as
endomembranas são raras, embora possam ser encontradas em muitas bactérias fotossintéticas.
PRODUÇÃO E UTILIZAÇÃO DE ENERGIA: Respiração celular é o processo de conversão das ligações
químicas de moléculas ricas em energia que poderão ser usadas nos processos vitais. Ela pode ser de dois
tipos, respiração anaeróbia (sem utilização de oxigênio) e respiração aeróbia (com utilização de oxigênio). A
respiração celular é o processo de obtenção de energia mais utilizado pelos seres vivos. Na respiração,
ocorre a libertação de dióxido de carbono, energia e água e o consumo de oxigênio e glicose. A organela
responsável por essa respiração é a mitocôndria na célula animal e o cloroplasto na célula vegetal. Do
ponto de vista da bioquímica, respiração celular é o processo de conversão das ligações químicas de
moléculas ricas em energia que possa ser usada nos processos vitais.
Glicólise: O processo básico da respiração celular é a quebra da glicose ou Glicólise, que se pode
expressar pela seguinte equação química:
C6H12O6 + 6O2 → 6CO2 + 6H2O + energia.
Obs.: A energia liberada pela respiração é o ATP (adenosina trifosfato), que pode perder um grupo fosfórico
e transformar-se em ADP (adenosina difosfato).
Fermentação e glicólise:
Semelhança básica:
Ambas permitem que a célula obtenha energia sempre a partir de combustíveis como a glicose.
Diferenças:
Na respiração é necessário o oxigênio, enquanto que na fermentação não é necessário.
Os resíduos da respiração são água e gás carbônico.
Os resíduos da fermentação são álcool e gás carbônico.
A equação química que representa a fermentação alcoólica é:
1C6H12O6 => 2C2H5OH + 2CO2 + ENERGIA
glicose => álcool etílico + gás carbônico + energia.
25
O NÚCLEO INTERFÁSICO: A interfase é o processo pelo qual a célula produz proteínas, aumenta de
tamanho e ocorre a duplicação do DNA presente na cromatina. É o período que antecede a divisão celular.
DIVISÃO CELULAR: Há dois tipos fundamentais de divisão celular: A mitose e a meiose.
Mitose: Uma célula mãe se divide originando duas células filhas idênticas à célula mãe. Trata-se de uma
célula diploide (com 46 cromossomos). A mitose compreende quatro fases ou etapas:
Prófase:
Os centríolos já duplicados afastam-se gradativamente para os pólos da célula, em torno dele
aparecem fibras que constituem o áster e entre os centríolos que se afastam formam-se as fibras
do fuso mitótico.
O núcleo desintegra distribuindo o seu material genético RNA ribossômico pela célula
Metáfase:
Os cromossomos atingem seu grau Máximo de espiralação, as cromátides de separam, tendo agora
cada uma delas um centrômero para constituir dois cromossomos-irmãos.
Anáfase:
Os cromossomos migram cada um para os pólos da célula.
Telófase:
Cada conjunto cromossômico atinge um dos pólos. Os cromossomos desespiralam gradativamente
e as duas novas cariotecas constituem-se a partir das membranas do retículo endoplasmático e
então ocorre a divisão do citoplasma com distribuição dos orgânulos entre as células filhas.
26
Meiose: Cada célula mãe origina quatro células-filhas, com metade do número de cromossomos que nela
existiam. São as células haplóides com apenas 23 cromossomos, células destinadas à reprodução.
Meiose I (Primeira Divisão Meiótica)
Prófase I – É a etapa mais marcante da meiose. Nela ocorre o pareamento dos cromossomos homólogos e
pode acontecer um fenômeno conhecido como crossing-over (também chamado de permuta). Como a
prófase I é longa, há uma sequência de eventos que, para efeito de estudo, pode ser dividida nas seguintes
etapas:
Inicia-se a espiralação cromossômica. É a fase de leptóteno (leptós = fino), em que os filamentos
cromossômicos são finos, pouco visíveis e já constituídos cada um por duas cromátides.
Começa a atração e o pareamento dos cromossomos homólogos; é um pareamento ponto por
ponto conhecido como sinapse (o prefixo sin provém do grego e significa união). Essa é a fase de
zigóteno (zygós = par).
27
A espiralação progrediu: agora, são bem visíveis as duas cromátides de cada homólogo pareado;
como existem, então, quatro cromátides, o conjunto forma uma tétrade ou par bivalente. Essa é a
fase de paquíteno (pakhús = espesso).
Ocorrem quebras casuais nas cromátides e uma troca de pedaços entre as cromátides homólogas,
fenômeno conhecido como crossing-over (ou permuta). Em seguida, os homólogos se afastam e
evidenciam-se entre eles algumas regiões que estão ainda em contato. Essas regiões são
conhecidas como quiasmas (qui corresponde à letra “x” em grego). Os quiasmas representam as
regiões em que houve as trocas de pedaços. Essa fase da prófase I é o diplóteno (diplós = duplo).
Os pares de cromátides fastam-se um pouco mais e os quiasmas parecem “escorregar” para as
extremidades; a espiralação dos cromossomos aumenta. È a última fase da prófase I, conhecida
por diacinese (dia = através; kinesis = movimento).
Enquanto acontecem esses eventos, os centríolos, que vieram duplicados da interfase, migram para os
polos opostos e organizam o fuso de divisão; os nucléolos desaparecem; a carioteca se desfaz após o
término da prófase I, prenunciando a ocorrência da metáfase I.
28
Metáfase I – os cromossomos homólogos pareados se dispõem na região mediana da célula; cada
cromossomo está preso a fibras de um só polo.
Anáfase I – o encurtamento das fibras do fuso separa os cromossomos homólogos, que são conduzidos
para polos opostos da célula, não há separação das cromátides-irmãs. Quando os cromossomos atingem os
polos, ocorre sua desespiralação, embora não obrigatória, mesmo porque a segunda etapa da meiose vem
a seguir. Às vezes, nem mesmo a carioteca se reconstitui.
Telófase I – no final desta fase, ocorre a citocinese, separando as duas células-filhas haploides. Segue-se
um curto intervalo a intercinese, que procede a prófase II.
Meiose II (segunda divisão meiótica)
Prófase II – cada uma das duas células-filhas tem apenas um lote de cromossomos duplicados. Nesta fase
os centríolos duplicam novamente e as células em que houve formação da carioteca, esta começa a se
desintegrar.
Metáfase II - como na mitose, os cromossomos prendem-se pelo centrômero às fibras do fuso, que partem
de ambos os polos.
Anáfase II – Ocorre duplicação dos centrômeros, só agora as cromátides-irmãs separam-se (lembrando a
mitose).
Telófase II e citocinese – com o término da telófase II reorganizam-se os núcleos. A citocinese separa as
quatro células-filhas haploides, isto é, sem cromossomos homólogos e com a metade do número de
cromossomos em relação à célula que iniciou a meiose.
29
GLOSSÁRIO
* Abiogênese: Teoria que postula o surgimento espontâneo da vida apartir da matéria bruta.
* Acrossomo: Vesícula existente na cabeça do espermatozóide, originada do sistema golgiense, rica em
enzimas que permitem a digestão dos envoltórios do óvulo.
* Arqueobactérias: As primeiras bactérias abundantes no planeta há bilhões de anos.
*Átomo: A menor porção de um elemento que pode entrar em combinação
* Biogênese: Teoria que propõe que toda vida provém de outra vida.
* Cariolinfa: Suco nuclear.
* Carioteca: Membrana que envolve a cromatina, nas células eucarióticas. O mesmo que membrana
nuclear.
*Cianobactérias: Organismo unicelular que realiza fotossíntese e apresenta pigmentos(coloração), como
clorofila, dentre outros exemplos como as algas azuis.
* Coacervado: Agregado de macromoléculas orgânicas, que pode ter sido um estágio na formação da vida
em nosso planeta.
* Conspícua: Notável, respeitável.
* Criacionismo: Criação divina( explicação aceita pelos criacionistas sobre a origem da vida, segundo a qual
a vida surgiu por intervenção de uma divindade ou força superior).
30
* Difusão facilitada: Tipo de transporte através da membrana plasmática, do local de maior concentração
para o de menor concentração, mediante a participação de uma proteína carregadora.
*Difusão simples: Passagem de partículas, eventualmente em solução, de um local de concentração maior
para o de concentração menor. Na célula, acontece através da membrana plasmática.
* Glicocálix: Revestimento de polissacarídeos existente na face externa da membrana plasmática das
células epiteliais.
* Glicólise: Etapa inicial da respiração celular, que ocorre no hialoplasma e envolve a degradação da glicose,
com produção de energia.
*Glicose: Monossacarídeo muito utilizado pelas células para obtenção de energia.
* Glucagon: Hormônio do pâncreas que aumenta a concentração de glicose no sangue.
* Invaginação: Dobramento de um órgão cavo sobre se mesmo, como um dedo de luva revirado.
* Lipoprotéico: Constituído de lipídios associados a proteínas.
*Molécula: Conjunto de dois ou mais átomos ligados que caracteriza determinada substância química.
* Muco: Substância viscosa segregada pelas glândulas mucosas.
* Nucleotídeos: Unidade constituinte das moléculas dos ácidos nucléicos, formada por uma pentose, um
fosfato e uma base hidrogenada.
* Panspermia: Doutrina segundo a qual os germes dos seres organizados acham-se espalhados por toda
parte, apenas aguardando que circunstâncias favoráveis lhes promovam o desenvolvimento.
* Paramécio: Protozoário ciliado comum nas águas doces paradas.
* Polimerização: Agregação de moléculas da mesma classe, formando moléculas maiores, sem
desprendimento de átomos.
* Polímero: Composto formado por polimerização
* Proteína: Molécula orgânica complexa, constituída por aminoácidos ligados entre se. As proteínas podem
ser construtoras da matéria viva.
REFERÊNCIAS BIBLIOGRÁFICAS:
http://www.brasilescola.com/biologia/celulas-procariontes.htm
Barros, Carlos Ciências Carlos Barros, Wilson Roberto Paulino, -- Ed. Reform. – São Paulo : ática,
2006.
Silva Júnior, Cesar da, 1934 – Biologia – volume1 – 1ª série – As Características da Vida, Biologia
Celular, Vírus: Entre Moléculas e Células, a origem da vida e histologia animal/César da Silva Júnior,
Sezar Sasson. – 8. Ed. – São Paulo: Saraiva, 2005.
www.sobiologia.com.br/conteudos/Corpo/Celula.
31