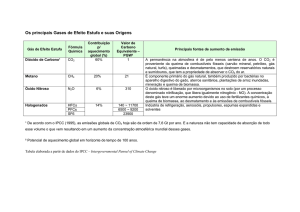
Revista Brasileira de Meteorologia, v.18, n.1, 87-96, 2003
SIMULAÇÕES DE FLUXO DE CARBONO EM UM
ECOSSISTEMA DE FLORESTA TROPICAL
SILVIA N. MONTEIRO SANTOS1 E MARCOS HEIL COSTA2
Departamento de Engenharia Agrícola, Universidade Federal de Viçosa UFV
Av. P. H. Rolfs, s/n, Viçosa-MG, 36571-000, Brasil; tel.: (31) 3899-1899; fax: (31) 3899-2735
1
E-mail: [email protected]; 2 Email: [email protected]
1
Endereço para correspondência.
RESUMO
Este trabalho usa o modelo SITE para simular o fluxo de CO2 numa floresta tropical amazônica. O
modelo SITE é um conceito simples baseado na micrometeorologia e dinâmica de ecossistemas
tropicais e seus principais processos físicos, químicos e biológicos de funcionamento. Uma
série de dados horários meteorológicos e de fluxos atmosféricos de CO2, coletados na Reserva
Florestal de Caxiuanã-PA, foram utilizados para calibrar e validar o modelo. O modelo reproduziu
satisfatoriamente os fluxos medidos de CO2, notavelmente algumas características de funcionalidade da vegetação, como a ocorrência do pico máximo do fluxo de CO2 antes das 12:00 e
oscilações no fluxo horário de CO2 devido a rápidas variações na nebulosidade. O padrão do
fluxo de CO2 é simulado no modelo principalmente pelas atividades metabólicas do ecossistema
(árvores e solo), sendo que as variações rápidas na turbulência ao longo do dia e da noite têm
uma importância secundária. Assim, a correta reprodução do fluxo horário de CO2 depende da
incorporação de conceitos de fisiologia vegetal e microbiologia de solos em modelos micrometeorológicos.
Palavras-chave: Floresta tropical, fluxo de carbono, micrometeorologia de ecossistemas.
ABSTRACT: SIMULATIONS OF CARBON FLUX IN A TROPICAL FOREST ECOSYSTEM
This work uses the model SITE to study the CO2 flux in an Amazonian tropical forest. SITE is a
simple model of micrometeorology and dynamics of tropical ecosystems, and is based on the
main physical, chemical and biological processes involved in the functioning of the tropical
ecosystems. Hourly meteorological and CO2 flux data, collected at the Forest Reserve of CaxiuanãPA, Brazil, were used to calibrate and validate the model. The model correctly reproduced the
measured fluxes of CO2, including characteristics that are relatively difficult to simulate, like the
timing of the maximum CO2 flux before 12:00, and oscillations in the hourly CO2 flux due to fast
variability in the cloud cover. CO2 flux is controlled mainly by the metabolical activities of the
ecosystem (canopy and soil), while the instantaneous micrometeorological characteristics are
secondary. Therefore, the correct reproduction of the hourly CO 2 flux depends on the
incorporation of plant physiology and soil microbiology concepts in micrometeorological models.
Key words: Tropical forest, CO2 flux, micrometeorology of ecosystems
1. INTRODUÇÃO
No balanço global de carbono, a queima de combustíveis fósseis e queimadas emitem quantidades consideráveis
de CO 2 para a atmosfera, parte do qual é dissolvido nos
oceanos, através da formação do íon HCO3-; e o restante
assimilado pela biosfera através do balanço entre a fotossíntese e a respiração das plantas e animais, ou permanece na
atmosfera, o que aumentou a sua concentração de 280 ppmv
no período pré-industrial para 360 ppmv na década de 1990.
A assimilação do carbono pela biosfera tem sido um
tema bastante discutido (Melillo et al., 1993; Ciais et al., 1995;
Schimel, 1995; Houghton et al., 1999). Basicamente,
argumenta-se que a biomassa da biosfera terrestre tem
crescido ao longo das últimas décadas em função do aumento
da eficiência fisiológica dos vegetais, do aumento da
concentração atmosférica de CO2 e também em função do
crescimento da vegetação natural de certas regiões devido
ao abandono de terras utilizadas na agricultura, do reflorestamento e de mudanças no manejo agrícola.
88
Simulações de Fluxo de Carbono em um Ecossistema de Floresta Tropical
A Amazônia é uma vasta região que apresenta diversos
ecossistemas, com destaque para a floresta tropical úmida,
um grande reservatório global de carbono. O debate sobre se
florestas primárias na Amazônia estão ou não atuando como
sumidouro de carbono está aberto, com posições consideravelmente conflitantes, havendo indicações de sumidouros
significativos e de estimativas próximas ao balanço (Grace et
al., 1995, 1996; Phillips et al., 1998; Houghton et al., 2000;
Richey et al., 2002; Miller et al., 2003).
O conhecimento atual da emissão/absorção de carbono pela floresta ainda é bastante limitado. O desenvolvimento de modelos que representam os processos ecológicos, biofísicos, biogeoquímicos e biogeográficos que
ocorrem em diferentes escalas de tempo permitiu que
diversos autores discutissem os prováveis efeitos da alteração da cobertura vegetal e da variabilidade do clima sobre a
dinâmica dos ecossistemas locais, incluindo o balanço de
carbono (Tian et al., 1998; Foley et al., 2002).
Entretanto, o estudo da fixação de carbono pelos
ecossistemas tropicais na maior parte das vezes tem sido
desconectado do contexto micrometeorológico, ou seja, do
contexto dos fluxos de gases e das suas causas. Assim, este
trabalho usa o SITE (Santos, 2001) como um simples modelo
de micrometeorologia e dinâmica de ecossistemas tropicais
para simular o fluxo de carbono num ecossistema de floresta
tropical amazônica. Espera-se assim descrever de maneira
mais detalhada os mecanismos que controlam os fluxos de
CO2 entre a floresta e a atmosfera.
2. METODOLOGIA
2.1. Descrição do Modelo
Para estudar o funcionamento dos ecossistemas
tropicais em relação às condições climáticas foi utilizado
neste trabalho um modelo simplificado de ecossistema
tropical de floresta (Simple Tropical Ecosystem Model
SITE), com a finalidade principal de simular os fluxos de
massa e energia entre o ecossistema e a atmosfera.
O modelo SITE foi desenvolvido através da simplificação de metodologias de modelagem previamente utilizadas,
visando maximizar a facilidade de uso do modelo e minimizar
o tempo de treinamento necessário à sua operação, mantendo
entretanto o alto rigor científico. Assim, o SITE se baseia
fundamentalmente em modelos previamente desenvolvidos,
principalmente o LSX (Pollard e Thompson, 1995), LSM
(Bonan, 1996), IBIS (Foley et al., 1996) e SiB2 (Sellers et al.,
1996), sendo muito mais simples. É um modelo dinâmico,
pontual, baseado no conceito big leaf (uma única camada),
que usa um intervalo de integração (dt) de uma hora, representando um ponto de terreno totalmente coberto por uma
floresta tropical perene. Pequenas modificações podem ser
necessárias para representar outros ecossistemas tropicais.
O modelo é estruturado com uma camada de dossel e duas
camadas no solo e é orientado ao uso em áreas instrumentadas com torres micrometeorológicas.
O modelo representa explicitamente diversos processos físicos, químicos e biológicos que ocorrem nos ecossistemas tropicais: balanço de radiação solar e infravermelha
no dossel, processos aerodinâmicos, fisiologia vegetal e
transpiração, transporte de massa e energia no dossel, fluxo
de calor e água no solo, e balanço de carbono no ecossistema.
O fluxo de radiação infravermelha é simulado considerando que a vegetação é um plano semitransparente com
uma emissividade dependente da densidade das folhas e
galhos. O módulo de radiação solar assume que toda reflexão
ocorre no topo do dossel, usando como dado de entrada o
albedo medido no topo da torre micrometeorológica. O
módulo aerodinâmico divide o dossel em três partes: entre o
topo do dossel e a atmosfera, assume um escoamento
predominantemente turbulento e um perfil logarítmico da
velocidade do vento com alto valor de rugosidade aerodinâmica; dentro da copa das árvores, assume um escoamento
difusivo e um perfil da velocidade do vento que diminui
exponencialmente dentro do dossel; abaixo da copa das
árvores, assume um novo perfil logarítmico do vento, com
baixo valor de rugosidade aerodinâmica. O módulo do
transporte de massa e energia no dossel são baseadas nos
princípios físicos da conservação de massa e energia. O
módulo do fluxo de água no solo considera a divisão do
solo em duas camadas, uma superficial que fornece água
para a evaporação livre e a outra com profundidade máxima
onde o sistema radicular consegue extrair água para a
transpiração. O fluxo de calor no solo é calculado em função
do balanço de energia á superfície e nas equações propostas
por Campbell e Norman (1998), e o módulo do balanço de
carbono no ecossistema é adaptada do modelo IBIS (Foley
et al., 1996), calculando o fluxo de carbono através de
equações prognósticas. Os componentes essenciais ao ciclo
do carbono (fisiologia vegetal e balanço de carbono) são
descritos em detalhe a seguir. Uma descrição detalhada de
todo o modelo pode ser obtida em Santos (2001).
2.2. Fisiologia Vegetal
Seria difícil determinar a taxa de CO2 absorvida pelas
folhas durante o processo da fotossíntese sem utilizar
conceitos de fisiologia vegetal. O fluxo líquido de CO 2
através do estômato é o principal determinante da condutância estomática, que por sua vez relaciona-se com a
evapotranspiração (fluxo de calor latente), importante em
estudos atmosféricos e hidrológicos. Este módulo do modelo
SITE se baseia nas equações propostas por Farquhar et al.
89
Silvia N. Monteiro Santos e Marcos Heil Costa
(1980), Collatz et al. (1991, 1992), Foley et al. (1996) e
Campbell e Norman (1998).
Os processos estomáticos são parametrizados por um
esquema conectado fotossíntese-condutância, que considera a difusão de CO2 entre a folha e o ar. Essa difusão do
CO2 no percurso atmosfera-cloroplastos não ocorre como
um gás e sim na fase líquida, pois em determinado momento
próximo as células, o CO2 é dissolvido em lâmina de água.
Com base nas equações de Collatz et al.(1991), o
modelo considera que a difusão do CO2 através do estômato
pode ser descrita por
An
gb
(01)
1, 6 A n
−
gs
(02)
CO 2s = CO 2a −
CO 2i = CO 2s
gs =
m A n h12
+ b
CO 2s
(03)
em que An é a taxa de fotossíntese líquida (mol CO2 m -2 s-1);
CO2a, CO2s e CO2i são as concentrações de CO2 na atmosfera;
na superfície da folha e nos espaços intercelulares das folhas
(mol mol-1); gb é a condutância da camada limite da folha e gs
é a condutância estomática (mol CO2 m-2 s-1); h12 é a umidade
relativa do ar na superfície foliar (igual à umidade relativa
média do ar dentro da copa do dossel); e m e b são coeficientes determinados empiricamente.
A taxa de fotossíntese líquida é o balanço entre os
processos de absorção (Ag) e de liberação de CO2 (Ru)
An = Ag − R u
(04)
em que Ag = min (Je, Jc) é a fotossíntese bruta, considerada
pelo modelo como sendo a taxa mínima limitante de duas
capacidades potenciais (Je e Jc), e Ru = γVmTm expresso em
função do coeficiente de respiração da folha da enzima
Rubisco, γ (Collatz et al., 1991) e da capacidade máxima da
enzima Rubisco, Vm = VmáxTvm, expressa a partir do parâmetro
Vmáx e atenuada pelos fatores de estresse hídrico do solo
(St) e de estresse de temperatura Tvm.
Este método para cálculo da fotossíntese bruta é uma
simplificação do método de Collatz et al. (1991), que prevê
três capacidades potenciais (Je, Jc, Js) e uma solução baseada
em uma equação quadrática. Entretanto, testes preliminares
indicaram que a capacidade potencial Js nunca era limitante
para concentrações atuais de CO2, e foi eliminada, simplicando a solução das equações.
A taxa de fotossíntese bruta limitada pela luz
(mol CO2 m-2 s-1) é expressa como:
J e = α3 APAR
CO 2i − Γ*
CO 2i + 2Γ*
(05)
[O 2 ]
é o ponto de compensação da luz (mol mol-1);
2τ
[O2] é a concentração de oxigênio (0,210 mol mol-1); α3 é a
eficiência quântica intrínseca para plantas C3; τ é a taxa do
parâmetro cinemático descrito a partir da enzima para
carboxilase ou oxigenase e APAR é a densidade de fluxo de
radiação fotossinteticamente ativa efetivamente absorvida
pela planta (mol fótons m-2 s-1).
A taxa de fotossíntese bruta limitada pela enzima
Rubisco (mol CO2 m-2 s-1) é dada por:
em que Γ* =
Jc =
Vm (CO 2i − Γ* )
[O 2 ]
CO 2i + K c 1 +
Ko
(06)
em que Kc e Ko são constantes de Michaelis, a primeira para
fixar o CO2 e a segunda para inibir o oxigênio, respectivamente. Os parâmetros fisiológicos utilizados no modelo estão
na Tabela 1, onde os valores de τ, Kc, Ko e Tvm foram obtidos
pelas respectivas equações adotadas e os demais valores
(Vmáx, b, m, α3, θ3 e γ) foram retirados da literatura e, eventualmente, foram otimizados durante o processo de calibração.
90
Simulações de Fluxo de Carbono em um Ecossistema de Floresta Tropical
2.3. Balanço de Carbono
A parametrização do balanço de carbono, adaptada
do modelo IBIS (Foley et al., 1996), calcula o fluxo de carbono
através de equações prognósticas. O carbono fixado pela
fotossíntese pode ser alocado em quatro diferentes reservatórios nas plantas: folhas, troncos e galhos, raízes finas e
raízes grossas. A quantidade de carbono alocada nesses
reservatórios é calculada usando equações diferenciais,
supondo uma fração fixa de alocação em cada reservatório e
tempos de residência fixos:
dCi
C
= a i NPP − i
τi
dt
(07)
em que i representa as folhas (i=u), galhos (i=s), raízes finas
(i=f) ou grossas (i=r), respectivamente.
O módulo de carbono no solo se baseia na decomposição de folhas e galhos por bactérias, em função da temperatura, umidade do solo e da massa de carbono na manta
orgânica, sendo semelhante ao usado no modelo IBIS,
porém, com algumas modificações.
dLi Ci
=
− f g g g h i Li
τi
dt
dDi Ci
=
−
τi
dt
(f
g
+ fd )
2
(08)
(g
g
+ gd )
2
h i Di
(09)
em que Li é a matéria orgânica morta da folha (i = u) ou galho
(i = s) sobre o solo, Di é a matéria orgânica morta das raízes
finas (i = f) e grossas (i = r) (kg C m-2), hi é a taxa de respiração
da manta orgânica de folhas (i = u), galhos (i = s), raízes
finas (i = f) ou grossas (i = r) (kg C kg -1 C s -1 );
(T − 283,16 ) 10 )
e g g = 0,25 + 0,75x i são funções de
f (Ti ) = 2( i
temperatura e umidade do solo na camada g (i = g) e camada
d (i = d) (adimensional). Os valores inicias da quantidade de
carbono em cada reservatório são mostrados na Tabela 2.
A produção líquida do ecossistema (NEP, kg C m-2 s-1)
é expressa pela diferença entre a respiração heterotrófica do
solo (Rsoil) e a produção primária líquida (NPP)
NEP = R soil − NPP
em que R soil = f g g g ( h u Lu + h s Ls ) +
(10)
(f
g
+ fd ) (gg + gd )
2
2
( h f Df + h r D r )
é calculado pela soma dos produtos das funções de
temperatura, umidade e taxa de decomposição para
c a d a reservatório de matéria orgânica no solo e
NPP = 0, 012 (1 − n ) ∫ ( A g − R aut
) dt
é expresso em
função da fotossíntese bruta, A g (mol CO 2 m -2 s -1) e da
respiração autotrófica do dossel (folhas, galhos e raízes). A
produtividade líquida do ecossistema é equivalente ao fluxo
total de CO2 entre a superfície e a atmosfera, similar à variável
medida em torres micrometeorológicas.
91
Silvia N. Monteiro Santos e Marcos Heil Costa
2.4. Descrição dos Dados e área Experimental
3. RESULTADOS E DISCUSSÃO
Uma série de dados de 28 dias, coletadas na estação
menos chuvosa do ano, no período de 29 de agosto a 25 de
setembro de 1999 (dia do ano 241-268), foi utilizada para a
calibração do modelo, e uma série de 41 dias, na estação
chuvosa, no período de 16 de abril a 27 de maio de 1999 (dia
do ano 106-147) foi utilizada para a validação do modelo.
Os dados utilizados na simulação do modelo fazem
parte da pesquisa realizada pelo projeto ECOBIOMA, parte
do Experimento de Grande Escala da BiosferaAtmosfera na
Amazônia (LBA). Os dados de entrada do modelo (forçantes
atmosféricas) são temperatura e umidade específica do ar,
velocidade horizontal do vento, radiação de onda curta
incidente, saldo de radiação de onda longa, albedo do dossel,
precipitação e pressão atmosférica, todos medidos acima do
dossel). Estes foram coletados em intervalos de 30 minutos
no período de abril a setembro de 1999, em um sítio
micrometeorológico na área experimental de floresta nativa
localizada na Reserva Florestal de Caxiuanã (01º4230S, 51º
3145W, 60 m), localizada no município de Melgaço-PA, a
cerca de 400 km a oeste da cidade de Belém-PA. A torre
micrometeorológica está instalada num ponto da reserva com
uma bordadura homogênea de pelo menos 3 km em todas as
direções.
O modelo usa como dados de entrada a radiação solar
incidente, albedo, radiação infravermelha incidente, temperatura e umidade do ar, precipitação e velocidade horizontal
do vento. As medições micrometeorológicas dos fluxos de
CO 2, água e energia foram obtidas através da técnica da
covariância dos vórtices turbulentos. O sistema contém um
anemômetro sônico tridimensional (Solent 1012R2) para medir
os três componentes do vento; um analisador de gás por
infravermelho (IRGA) LI-6262 (closed-path), para medições
de concentrações de CO2 e H2O; um microcomputador portátil
e um software Edisol (Universidade de Edinburgh) para
aquisição e análise dos dados. As medições das variáveis
meteorológicas (temperatura do bulbo seco e úmido, radiação
incidente e refletida de onda longa e curta, velocidade e
direção do vento, e precipitação) foram obtidos através de
uma estação meteorológica automática, registrando média
de 30 minutos e armazenadas em um sistema de aquisição de
dados Campbell Scientific. Todas essas medições foram
coletadas no topo de uma torre de alumínio, a 56 m de altura
(aproximadamente 16 m acima da copa do dossel). A
concentração de CO2 foi medida ao longo do perfil do dossel,
para transformação das medições de fluxo de CO2 em valores
de NEP. Inicialmente, o fluxo de calor no solo foi medido
também, mas devido a um defeito no sensor, essas medições
foram abandonadas, o que não nos permitiu fechar o balanço
de energia no ecossistema.
Os valores médios simulados de NEP para os períodos
de 17 de abril (107) a 27 de maio (147), utilizado na validação,
e 29 de agosto (241) a 25 de setembro (268) de 1999, utilizado
na calibração do modelo foram de -0,79 e -0,90 kg C ha-1 h-1,
respectivamente, apresentando um erro de -6,5% em relação
ao observado (fluxo turbulento + variação no armazenamento)
no período da validação e 6,8% em relação ao observado no
período de calibração. Na convenção usada neste trabalho,
valores negativos de NEP equivalem à assimilação de carbono pelo ecossistema, enquanto os positivos significam
emissão.
Carswell et al. (2002) encontraram um valor observado
de -0,61 kg C ha-1 h-1 para o local em estudo, com base num
período mais longo de 16 de abril a 19 de outubro de 1999;
Malhi et al. (1998) encontraram um NEP de -0,65 kg C ha-1 h-1
para Cuieiras, Manaus, no período de 13 a 21 de novembro
de 1995. Esses valores observados foram calculados usando
todos os valores disponíveis da série de dados de fluxo de
CO2.
Tem-se argumentado, entretanto, que o transporte de
CO2 em noites calmas (sem turbulência) ocorre por meio de
processos não medidos pelo sistema de correlação de
vórtices turbulentos (Goulden et al., 1996). Por exemplo, Miller
et al. (2003) encontraram uma dependência do NEP noturno
com a turbulência acima do dossel, mostrando que o NEP
noturno era reduzido para valores de velocidade de fricção
do vento no perfil acima do dossel (u*) menores do que
0,2 m s -1, ao mesmo tempo em que as medições de fluxo de
CO 2 do solo não indicavam que a respiração do solo
estivesse relacionada com a turbulência acima do dossel.
Assim, conclui-se que o sistema de correlação de
vórtices turbulentos não fornece valores confiáveis quando
não há turbulência. O limite de funcionamente destes
sistemas ainda é objeto de debate, mas em algumas comparações neste trabalho estamos colocando em dúvida e filtrando
(desprezando) os valores de fluxos medidos durante a noite
quando a velocidade de fricção é inferior a 0,2 m s-1. Assim,
por exemplo, enquanto o RMSE (raiz do erro quadrado médio)
ao se comparar a série de dados simulados com todos os
dados observados é 0,174 kg C ha -1 h -1, o RMSE ao se
comparar a série de dados simulados com os dados observados filtrados é de apenas 0,15 kg C ha-1 h -1. No restante
desta discussão, os resultados apresentados geralmente não
consideram eliminação de dados da série, a não ser quando
especificado.
A Figura 1 mostra o comportamento dos processos
associados aos fluxos de CO2, e o NEP simulado e observado
nos períodos de 19 a 26 de abril (109 a 116). A fotossíntese
bruta (Ag) ou Produtividade Primária Bruta (GPP) é o mínimo
92
Simulações de Fluxo de Carbono em um Ecossistema de Floresta Tropical
entre a taxa de fotossíntese limitada pela disponibilidade de
PAR (Je) e a taxa de fotossíntese limitada pela atividade da
enzima rubisco (Jc) (Figura 1a). Notase valores nulos de Je
durante a noite e máximos próximos das 12:00 horas, horário
local. Por outro lado, Jc apresenta um valor mínimo durante a
tarde, o que é devido principalmente à dinâmica de carbono
através dos estômatos, que faz que CO2i seja maior durante
a noite do que durante o dia. A dependência inversa de Kc e
K o com a temperatura também contribuem, de maneira
secundária, para o formato da curva de Jc.
A Figura 1b mostra a produtividade primária líquida
(NPP) da floresta, ou seja, a diferença entre a fotossíntese
bruta e a respiração autotrófica de folhas, raízes finas e da
parte viva de troncos, galhos e raízes grossas. Basicamente,
o NPP segue o mesmo comportamento de Ag, com valores
um pouco inferiores. Pode-se examinar também nestas figuras
o comportamento da respiração heterotrófica do solo (Rsoil).
Nota-se, nos picos durante a tarde, a dependência de Rsoil
com a temperatura do solo. A diferença entre a respiração
do solo (Rsoil) e o NPP é a troca líquida do ecossistema (Figura
1c). Apesar do modelo reproduzir o comportamento geral do
fluxo de CO 2, algumas variações em escala horária não
conseguem ser capturadas. Esses picos de CO2 geralmente
ocorrem em horário de forte velocidade do vento, e representam os momentos em que a condutância aerodinâmica é mais
importante para definir o fluxo de CO2 do que as atividades
metabólicas do ecossistema.
A Figura 1 mostra que a taxa de fotossíntese aumenta
rapidamente a partir do amanhecer. Na segunda metade da
manhã, a atividade da enzima Rubisco passa a limitar a
atividade fotossintética, que atinge o seu pico antes das
12:00, hora local. Os valores de pico de NEP simulados
oscilaram entre -8 a -9 kg C ha-1 h-1. Carswell et al. (2002)
obtiveram um valor máximo médio de -8,2 kg C ha-1 h-1 para a
floresta de Caxiuanã no período de 108 a 114 para o mesmo
ano em estudo, com pico médio às 11:00, hora local. Malhi
et al. (1998) constataram um pico de -7,8 kg C ha-1 h-1 em
Cuieiras, na Amazônia central.
As condições médias próximas ao meio-dia incluem a
radiação solar mais intensa e temperaturas do ar e da folha
mais elevadas. Essas condições contribuem para um ligeiro
fechamento dos estômatos e diminuição da fotossíntese,
evitando a perda excessiva de água por evaporação e
diminuindo as taxas de assimilação de CO2. Pode-se observar
que justamente nesse horário, a fotossíntese bruta deixa de
ser controlada pela disponibilidade de radiação, passando a
ser limitada pela ação da enzima Rubisco durante a fase
escura. Na transição da fase clara para a fase escura ocorre
a assimilação máxima de CO2. No fim da tarde, a assimilação
de CO 2 pela planta passa novamente a ser limitada pela
disponibilidade de radiação solar (Figura 1a).
Verifica-se ainda na Figura 1a que o GPP apresentou
um valor médio de aproximadamente 4,3 kg C ha-1 h-1 em todos
os períodos. Carswell et al. (2002) calcularam um valor de
GPP de aproximadamente 4,1 kg C ha-1 h-1 para a floresta de
Caxiuanã. O GPP encontrado por Malhi et al. (1998) foi de 3,5
kg C ha-1 h-1 em Cuieiras, Manaus, enquanto Lloyd et al.
(1995) obtiveram um GPP em torno de 2,7 kg C ha-1 h-1 para a
Reserva Florestal de Jaru, Rondônia.
Na Figura 1b verificase que o NPP apresentou picos
em torno de 11 kg C ha-1 h-1 e valor médio acumulativo de
aproximadamente 2,3 kg C ha-1 h-1 em todos os períodos. O
valor simulado pelo modelo é semelhante ao simulado por
Kucharik et al. (2000), que obtiveram valores médios de 2,2
kg C ha-1 h-1 para as florestas tropicais e maior que o verificado por Melillo et al. (1993) que estimaram valores médios de
NPP de 1,3 kg C ha-1 h-1 para as florestas tropicais.
A respiração heterotrófica do solo simulada
apresentou valor máximo de aproximadamente 1,7 kg C ha-1 h1
e valor médio estimado em torno de 1,5 kg C ha-1 h-1 no
período considerado. Esses valores simulados estão próximos dos valores médios obtidos por Trumbore et al. (1995) e
Davidson et al. (2000) de 1,71 e 2,3 kg C ha-1 h-1, respectivamente, na fazenda Victoria, no município de Paragominas-PA. A
maior parte das referências disponíveis na literatura dizem
respeito à respiração total (autotrófica e heterotrófica) do solo.
Carswell et al. (2002) constataram uma variação sazonal para a
respiração noturna de 3,1 a 4,0 kg C ha-1 h-1, para a Reserva
Florestal de Caxiuanã. Rocha et al (1996) encontraram um valor
médio de aproximadamente 2,7 kg C ha-1 h-1 na Reserva Ducke,
correspondente ao período de setembro de 1983 a agosto 1985,
tendo também constatado que o solo contribui com cerca de 70
a 80% do total de carbono emitido para a atmosfera. Malhi et al.
(1998) obtiveram um valor médio em torno de 2,8 kg C ha-1 h-1
para a respiração do solo em Cuieiras, Manaus. Grace et al.
(1996) encontraram valores de respiração noturna que variaram
de 2,6 a 3,0 kg C ha-1 h-1 na Reserva Jaru, Rondônia.
Esta análise de modelagem também reproduziu corretamente o horário em que o fluxo máximo de CO2 ocorre, 11:00
neste caso. Modelos que simulam a fotossíntese baseados
apenas na radiação solar geralmente simulam o fluxo máximo
de CO2 ocorrendo às 12:00. Para obter a correta representação
do horário do fluxo máximo de CO2, é necessário considerar
que a fotossíntese bruta é limitada não apenas pela radiação
solar, mas também pela atividade enzimática.
A Figura 2 é semelhante à Figura 1, porém destaca o
funcionamento do modelo em dias com nebulosidade.
Nesses dias (por exemplo, 125 e 129), a radiação solar é
reduzida pela presença de nuvens, e J e passa a limitar a
fotossíntese bruta. As reduções de fixação de CO2 observadas na torre são acompanhadas pelo modelo, mesmo no caso
de variações rápidas na nebulosidade, como no dia 129.
93
Silvia N. Monteiro Santos e Marcos Heil Costa
Na Figura 3 observa-se a dispersão do fluxo de CO2
para um total de 37 dias, no período de 16 de abril (107) a 27
de maio (147) de 1999. Para a construção desse gráfico, foram
eliminados os valores medidos e simulados de fluxo de CO2
quando o escoamento não era turbulento (u* < 0,2 m s-1).
Apesar da boa correlação entre os valores simulados e
observados, nota-se, na Figura 2, uma incapacidade do
modelo em simular valores elevados de fluxo de CO2 que
ocorrem durante a noite. Essa limitação está provavelmente
Figura 2. Igual à Figura 1, porém para o período de 4 a 9 de maio de
(124 a 129) de 1999.
Figura 1. Comportamento dos processos associados ao fluxo de
CO 2 observado e simulado. (a) fotossíntese bruta (A g) e seus
componentes Je e Jc; (b) respiração heterotrófica do solo (Rsoil) e
produção primária líquida (NPP) e (c) troca líquida de ecossistema
(NEP) no período de 19 a 26 de abril (109 a 116) de 1999, na
floresta de Caxiuanã. O sinal negativo corresponde ao acúmulo de
CO2 no ecossistema.
Figura 3. Dispersão do Fluxo de CO2 para o período de 16 de abril
(107) a 27 de maio (147) de 1999.
94
Simulações de Fluxo de Carbono em um Ecossistema de Floresta Tropical
associada à representação simplificada dos processos de
respiração heterotrófica no solo.
A Figura 4 mostra o fluxo acumulado de CO2 neste
ecossistema (observado e simulado), a partir do dia 16 de
abril de 1999 (dia 106). Pelo comportamento das duas curvas,
verifica-se que o modelo não tem tendência dominante nem
de subestimar nem de sobreestimar os valores observados.
Deve-se notar que a umidade do solo praticamente
não influenciou os fluxos simulados de CO2, nem na estação
chuvosa e nem na estação seca, ou seja, em nenhum dos dois
períodos estudados a floresta esteve sob estresse hídrico. Na
verdade, tanto nos dados observados quanto nos simulados,
a assimilação de CO2 foi ligeiramente maior na estação seca
do que na estação chuvosa, provavelmente devido à maior
incidência de radiação solar devido à menor nebulosidade.
• A correta reprodução do fluxo horário de CO2 depende
da incorporação de conceitos de fisiologia vegetal e
microbiologia de solos em modelos micrometeorológicos.
Devido à sua facilidade de uso, o modelo SITE seria
apropriado para utilização em disciplinas de micrometeorologia em cursos de pós-graduação, bem como em pesquisas
simples na área de micrometeorologia de ecossistemas.
5. AGRADECIMENTOS
Os autores agradecem ao suporte financeiro dado pela
CAPES, CNPq (programa PADCT) e WWF (Fundo Mundial
para a Natureza), para a efetiva realização deste trabalho.
6. REFERÊNCIAS BIBLIOGRÁFICAS
BONAN, G.B. A Land surface model (LSM version 1.0) for
ecological, hydrological, and atmospheric studies:
technical description and users guide. NCAR Technical
note TN-417+STR, 1996. 150p.
CAMPBELL, G.S.; NORMAN, J.M. An introduction to
environmental biophysics. 2. ed. New York: SpringerVerlag, 1998. 286p.
Figura 4. Comportamento cumulativo total do Fluxo de CO2 para
37 dias, no período de 16 de abril (106) a 22 de maio (106 a 142)
de 1999.
4. CONCLUSÕES E RECOMENDAÇÕES
Este trabalho utilizou o modelo SITE para estimar os
fluxos de carbono no ecossistema de floresta tropical. A
análise dos resultados obtidos através de diversas simulações para o ecossistema de floresta tropical amazônica
permitiram as seguintes conclusões:
• Os valores de fluxo de CO2 simulados pelo modelo, na
maioria dos casos, se ajustaram adequadamente aos
dados observados na área experimental. Destaca-se a
reprodução de detalhes relativamente difíceis de serem
simulados, como o pico do NEP ocorrendo antes das
12:00, e oscilações no fluxo horário de CO2 devido a
rápidas variações na nebulosidade.
• O padrão do ciclo diurno do fluxo total de CO2 é
controlado no modelo principalmente pelas atividades
metabólicas do ecossistema (árvores e solo). As
variações rápidas na turbulência ao longo do dia e da
noite têm uma importância secundária.
CARSWELL, F. E.; COSTA, A.C.L.; PALHETA, M.; MALHI,
Y.; MEIR, P.; COSTA, J.P.R.; RUIVO, M.L.; LEAL, L.S.M.;
COSTA, J.M.N.; CLEMENTE, R.J.; GRACE, J. Seasonality
in CO2 and H2O flux at an eastern Amazonian Rain Forest.
J. Geophys. Res.- Atmos., v.107, n.D20, art. n. 8076, 2002.
CIAIS, P.; TANS, P.P.; TROLIER, M.; WHITE, J.W.C.;
FRANCEY, R.J. A large northern hemisphere terrestrial
CO 2 sink indicated by 13 C/ 12 C of atmospheric CO 2 .
Science, v.269, p.1098-1102, 1995.
COLLATZ, G.J.; BALL, J.T.; GRIVET, C.; BERRY, J.A.
Physiological and environmental regulation of stomatal
conductance, photosynthesis and transpiration: a model
that includes a laminar boundary layer. Agr. Forest.
Meteorol., v.54, p.107-136, 1991.
COLLATZ, G.J.; CARBO, M.R.; BERRY, J.A. Coupled
photosynthesis-stomatal conductance model for leaves of
C4 plants. Aust. J. Plant Physiol., v.19, n.5, p.519-538, 1992.
DAVIDSON, E.A.; VERCHOT, L.V.; CATTÂNIO, J.H.;
ACKERMAN, I.L.; CARVALHO, J.E.M. Effects of soil water
content on soil respiration in forests and cattle pastures of
eastern Amazonia. Biogeochemistry, v.48, p.53-69, 2000.
95
Silvia N. Monteiro Santos e Marcos Heil Costa
FARQUHAR, G.D.; CAEMMERER, S.V.; BERRY, J.A. A
biochemical model of photosynthetic CO2 assimilation in
leaves of C3 species. Planta, v.149, p.78-90, 1980.
LEUNING, R. A critical appraisal of a combined stomatalphotosynthesis model for C3 plants. Plant, Cell and
Environment, v.18, p.339-355, 1995.
FOLEY, J.A.; PRENTICE, I.C.; RAMANKUTTY, N.; LEVIS,
S.; POLLARD, D.; SITCH, S.; HAXELTINE, A. An
integrated biosphere model of land surface processes,
terrestrial carbon balance and vegetation dynamics.
Global Biogeochemical Cycles, v.10, p.603-628, 1996.
LLOYD, J.; GRACE, J.; MIRANDA, A.C.; MEIR, P.;
WONG, S.C.; MIRANDA, H.S.; WRIGHT, I.R.; GASH,
J.H.C.; MCINTYRE, J. A simple calibrated model of
Amazon rainforest productivity based on leaf biochemical
properties. Plant, Cell and Environment, v.18, p.11291145, 1995.
FOLEY, J.A.; BOTTA, A.; COE, M.T.; COSTA, M.H. The El
Niño/Southern Oscillation and the climate, ecosystems
and rivers of Amazonia. Global Biogeochemical Cycles.
v.16., n.4, 2002. 1132, doi: 10.1029/2002 6B 001872.
GOULDEN, M.L.; MUNGER, J.W.; FAN, S.-M.; DAUBE, B.C.;
WOFSY, S.C. Measurements of carbon sequestration by
long-term eddy covariance: Methods and a critical
evaluation of accuracy. Global Change Biology, v. 2, n.3,
p.169-182, 1996.
GRACE, J.; LLOYD, J.; MCINTYRE, J.; MIRANDA, A.C.;
MEIR, P.; MIRANDA, H.S.; NOBRE, C; MONCRIEFF, J.;
MASSHEDER, J.; MALHI, Y.; WRIGHT, I.; GASH, J.
Carbon dioxide uptake by an undisturbed tropical rain
forest in Southwest Amazonia, 1992 to 1993. Science,
v.270, p.778780, 1995.
GRACE, J.; LLOYD, J.; MCINTYRE, J.; MIRANDA, A.C.;
MEIR, P. Carbon dioxide flux over Amazon rainforest in
Rondônia. In: GASH, J.H.C.; NOBRE, C.A; ROBERTS,
J.M.; VICTORIA, R.L. (Eds.). Amazonian deforestation
and climate. Chichester: John Wiley and Sons, 1996.
p.307-318.
HOUGHTON, R.A.; HACKLER, J.L.; LAWRENCE, K.T. The
U.S. Carbon budget: contributions from landuse change.
Science, v.285, p.574-578, 1999.
HOUGHTON, R. A.; SKOLE, D. L.; NOBRE, C. A.; HACKLER,
J. L.; LAWRENCE, K. T. & CHOMENTOWSKI, W. H.
Annual fluxes of carbon from deforestation and regrowth
in the Brazilian Amazon. Nature, v.403, p.301304, 2000.
KUCHARIK, C.J.; FOLEY, J.A.; DELIRE, C.; FISHER, V.A.;
COE, M.T.; GOWER, S.T.; LENTERS, J.D.;
YOUNGMOLLING, C.; NORMAN, J.M.; RAMANKUTTY,
N. Testing the performance of a dynamic global ecosystem
model: water balance, carbon balance end vegetation
structure. Global Biogeochemical Cycles, v.14, p.795825, 2000.
MALHI, Y.; NOBRE, A.D.; GRACE, J.; KRUIJT, B.;
PEREIRA, A.C.; SCOTT, S. Carbon dioxide transfer over
a central Amazonian rain forest. J. Geophys. Res., v.31,
p.31593-31612, 1998.
MELILLO, J.M.; MCGUIRE, A.D.; KICKLIGHTER, D.W.;
MOORE III, B.; VÖRÖSMARTY, C.J.; SCHLOSS, A.L.
Global climate change and terrestrial net primary
production. Nature, v.363, p.234-240, 1993.
MILLER, S.D.; GOULDEN, M.L.; MENTON, M.C.;
ROCHA, H.R. DA; FREITAS, H.C. Annual CO 2
exchange by a tropical Forest. Ecological Applications.
2003.
PHILLIPS, O.L.; MALHI, Y.; HIGUCHI, N.; LAURANCE, W.
F.; NÚÑEZ, P.V.; VÁSQUEZ, R.M.; LAURANCE, S.G.;
FERREIRA, L.V.; STERN, M.; BROWN, S.; GRACE, J.
Changes in the carbon balance of tropical forests:
evidence from longterm plots. Science, v.282, p.439-442,
1998.
POLLARD, D.; THOMPSON, S.L. The effect of doubling
stomatal resistance in a global climate model. Global and
Planetary Change, v.10, p.129-161, 1995.
RICHEY, J.E.; MELACK, J.M.; AUFDENKAMPE, A.K.;
BALLESTER, V.M.; HESS, L.L. Outgassing from
Amazonian Rivers and Wetlands as a Large Tropical
Source of Atmospheric CO2. Nature, v.416, p.617-620,
2002.
R O C H A , H . R . ; S E L L E R S , P. J . ; C O L L AT Z , G . J . ;
WRIGHT, I.R.; GRACE, J. Calibration and use of
the SiB2 model to estimate water vapor and carbon
exchange at the ABRACOS forest sites. In: GASH,
J.H.C.; NOBRE, C.A.; ROBERTS, J.M.; VICTORIA,
R.L. (Eds.). Amazonian Deforestation and
Climate. Chichester : John Wiley and Sons, 1996.
p.460-471.
96
Simulações de Fluxo de Carbono em um Ecossistema de Floresta Tropical
SANTOS, S.N.M. Modelo de fluxos de energia, água e CO2
aplicado em ecossistema de floresta tropical. ViçosaMG, 2001. 80p. Dissertação (Mestrado) - Universidade
Federal de Viçosa.
SCHIMEL, D.S. Terrestrial ecosystems and the carbon cycle.
Global Change Biology, v.1, p.77-91, 1995.
SELLERS, P.J.; RANDALL, D.A.; COLLATZ, G.J.; BERRY,
J.A.; FIELD, C.B.; DAZLICH, D.A.; ZHANG, C.;
COLLELO, G.D.; BOUNOUA, L. A revised land surface
parameterization (SiB2) for atmospheric GCMs .1. Model
formulation. Journal of Climate, v.9, p.676-705, 1996.
TIAN, H.; MELILLO, J.M.; KICKLIGHTER, D.W.; MCGUIRE,
A.D.; HELFRICH III, J.V.K.; MOORE III, B.;
VÖRÖSMARTY, C.J. Effect of interannual climate
variability on carbon storage in amazonian ecosystems.
Nature, v.396, p.664-667, 1998.
TRUMBORE, S.E.; DAVIDSON, E.A.; CAMARGO, P.B.;
NEPSTAD, D.C.; MARTINELLI, L.A. Belowground
cycling of carbon in forests and pastures of eastern
Amazonia. Global Biogeochemical Cycles, v.9, p.515528, 1995.