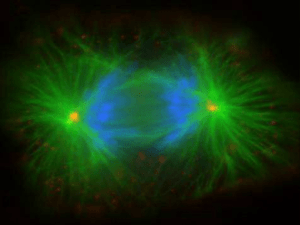
A célula em divisão
Mitose
Todas as células obedecem a um ciclo celular, período que compreende uma fase em que a célula
cresce e reproduz suas estruturas internas e outra durante a qual ela se divide.
O período da intérfase compreende três etapas (G1, S, G2) e algumas células podem permanecer
para sempre em G1, sem passar à fase de divisão, chamada M. Neste caso a etapa é chamada de G0,
como observado nos neurônios.
O ciclo celular dura, em média, de 12 a 24 horas. A fase M dura cerca de 1 hora. Esse é um valor
médio, havendo, naturalmente, muitas variações. Cada fase do ciclo celular é disparada por Cdks. As
ciclinas, como já sabemos, são proteínas que, uma vez ativadas, deflagram eventos específicos. A divisão
celular ou mitose é disparada pela M-Cdk.
A mitose é um evento bastante complexo, que tem por objetivo distribuir equitativamente o material
genético, duplicado na fase S, entre as duas células-filhas.
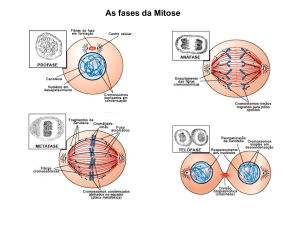
A mitose inclui uma sequência de eventos que, embora nem sempre possam ser claramente
delimitados, foram divididos em cinco fases: Prófase, Prometáfase, Metáfase, Anáfase e Telófase.
Uma célula em divisão é bastante diferente de uma célula em intérfase em, pelo menos, três
características:
• O envoltório nuclear, presente na célula interfásica, desaparece durante a divisão.
• Os cromossomos, que formam uma massa na célula interfásica, se condensam (espiralizam) e se
individualizam durante a divisão.
• Durante a divisão, os microtúbulos se rearranjam, dando origem ao fuso acromático.
As condições necessárias para que a célula entre na fase M são providenciadas durante a intérfase.
Nesse período os cromossomos se duplicam, permanecendo unidos pela região chamada centrômero.
Também se duplicam nesta fase os centrossomos. Eles
são o que chamamos centro organizador de microtúbulos, isto é,
todos os microtúbulos de uma célula partem daí. Nas células
animais, os centrossomos incluem um par de centríolos, cuja
estrutura formada por nove trios de microtúbulos é bem
característica. Observe essa estrutura na figura ao lado.
Os centríolos de um par não são idênticos. Um deles é dominante e possui filamentos que o
conectam à matriz pericentriolar (de onde partem os microtúbulos). Quando os centríolos de um par vão
se duplicar, eles se separam e cada um dá origem (nucleia) a um novo centríolo. Os dois pares de
centríolos permanecem próximos até o início da prófase.
Uma vez que os cromossomos e centrossomos estejam duplicados, a mitose pode ter início.
PRÓFASE
No início da prófase, o envoltório nuclear ainda se encontra intacto. É nesta etapa que os
cromossomos duplicados se condensam (espiralizam) e assumem sua forma característica de dois bastões,
as cromátides-irmãs, ligados pelo centrômero.
Como os cromossomos interfásicos, longos e finos, se enovelam dessa forma? A resposta está
numa proteína que possui dois domínios capazes de se ligarem à hélice de DNA. Por ser capaz de
promover a condensação dos cromossomos, à custa da hidrólise de ATP, foi denominada condensina. A
condensina funciona como uma pinça em que cada extremidade se liga a um ponto da cadeia de DNA e se
fecha em seguida, aproximando as duas. Na região do centrômero, uma proteína da mesma família mantém
as duas cromátides coesas. Seu nome? Coesina.
Embora o envoltório nuclear ainda esteja intacto, a migração dos centrossomos para os polos
opostos tem início nessa etapa. À medida que migram para polos opostos da célula, os microtúbulos se
irradiam a partir dos centrossomos. Por lembrar uma estrela, cada uma dessas estruturas é chamada áster.
Este é o início da formação do fuso acromático, que só estará completamente formado na etapa seguinte.
PROMETÁFASE
Dizia-se que nessa fase do ciclo celular o envoltório nuclear desaparecia. Na verdade, o envoltório
nuclear, o retículo endoplasmático e o complexo de Golgi não são visíveis nessa fase porque se
fragmentam em vesículas, permitindo que alguns microtúbulos do fuso acromático se liguem ao cinetócoro
dos cromossomos, já totalmente condensados. O cinetócoro é um complexo de proteínas que se liga aos
cromossomos na região do centrômero.
Os cromossomos só se ligam ao fuso pelo cinetócoro. Essa ligação parece ser feita de modo
aleatório, pelo sistema de tentativa e erro, isto é, os vários microtúbulos, que partem dos centrossomos de
acordo com a instabilidade dinâmica que lhes é característica, crescem e encolhem rapidamente.
Eventualmente, alguns deles tocam os cromossomos. Se esse contato acontecer na região certa, isto é, a
extremidade do microtúbulo fazendo contato com o cinetócoro, o cromossomo permanecerá ancorado
àquele microtúbulo. Como os cromossomos estão duplicados, os cinetócoros de cada uma das cromátidesirmãs ficarão ligados a microtúbulos de um polo. Os microtúbulos mais longos são mais fortes, puxando o
cromossomo na direção do centrossomo no qual têm sua origem. Estabelece-se, assim, um verdadeiro
cabo de guerra entre os dois polos. Essa disputa termina empatada, com os cromossomos alinhados no
equador da célula. Esse movimento dos cromossomos para o equador celular foi chamado de metacinese.
Quando todos os cromossomos se alinham de forma equidistante dos polos, significa que a divisão
celular entrou na metáfase.
Modernamente os eventos da prometáfase foram incorporados à prófase e o nome desta etapa foi
excluído da mitose.
METÁFASE
Na metáfase, o fuso acromático já está plenamente desenvolvido e os cromossomos se alinham no
plano equatorial da célula, equidistantes dos dois polos, formando a placa equatorial ou metafásica. Além
dos microtúbulos astrais – isto é, que compõem o áster –, e dos microtúbulos cinetocoriais – que se ligam
ao cinetócoro dos cromossomos –, o fuso possui microtúbulos que desempenham uma terceira função.
Trata-se de microtúbulos que partem dos polos opostos e se interpenetram no plano equatorial da célula.
Podemos chamá-los microtúbulos interpenetrantes. Os três tipos de microtúbulos são altamente
dinâmicos, polimerizando-se e despolimerizando-se mais rapidamente que os microtúbulos das células
interfásicas e formam o fuso acromático ou fuso mitótico. Conforme as forças exercidas pelos
microtúbulos cinetocoriais ligados a cada uma das cromátides-irmãs se equilibram, encolhendo de um lado
e alongando-se do outro, os cromossomos vão se dispondo na placa equatorial. Esta é a fase mais
demorada da mitose e a divisão só prossegue quando todos os cromossomos se encontram alinhados.
ANÁFASE
Na anáfase, as cromátides-irmãs se separam e cada uma delas é puxada pelos microtúbulos
cinetocoriais em direção a polos opostos. Como as cromátides-irmãs se separam se elas estão ligadas pela
coesina? A resposta é bastante simples: a enzima separase se encarrega de cortar as pontes formadas
pela coesina. Assim, cada cromátide é puxada para um polo pelos microtúbulos cinetocoriais respectivos. A
migração dos cromossomos para polos opostos resulta não apenas do encolhimento dos microtúbulos
cinetocoriais. Proteínas motoras associadas aos microtúbulos interpenetrantes também contribuem para
que haja entre eles um deslizamento que aumenta a distância entre os polos. O distanciamento provocado
pelo deslizamento entre microtúbulos do fuso é o prenúncio da última fase da divisão celular, a telófase.
TELÓFASE
A telófase corresponde ao final da mitose. Nessa etapa, os cromossomos já se encontram
segregados em polos opostos da célula e distanciados pelos microtúbulos que se superpõem. Esse
distanciamento é fundamental para que ocorra a distribuição do citoplasma e das organelas entre as duas
células-filhas. Esse processo é chamado citocinese.
Os microtúbulos astrais também exercem força de separação
entre as células-filhas deslizando sobre proteínas motoras ligadas à
membrana plasmática. A separação final entre as células-filhas depende
de um anel de constrição (ou anel contrátil) formado por filamentos de
actina e moléculas de miosina. Veja detalhe na figura ao lado.
Nas células animais a citocinese é centrípeta (de fora para
dentro) e nas células vegetais é centrífuga (de dentro para fora), pela
formação da lamela média.
Na telófase, a carioteca e os nucléolos reaparecem, e os
cromossomos desespiralizam e desindividualizam, voltando a compor a
cromatina nuclear.




![Ciclo celular [Modo de Compatibilidade]](http://s1.studylibpt.com/store/data/004234212_1-04dedc414d2f87755e3978361fad4b55-300x300.png)