CAPÍTULO 7 - Nervos cranianos e seus núcleos
Núcleos dos nervos cranianos aferentes
Núcleos dos nervos cranianos eferentes
Nervos cranianos
III: Nervo oculomotor
IV: Nervo troclear
VI: Nervo abducente
V: Nervo trigêmeo
VII: Nervo facial
VIII: Nervo vestibulococlear
IX: Nervo glossofaríngeo
X: Nervo vago
XI: Nervo acessório
XII: Nervo hipoglosso
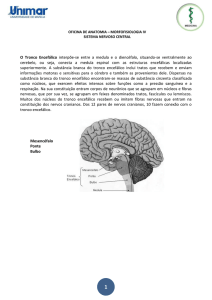
Fig. 7.1 A base do encéfalo, mostrando a localizaç o dos nervos cranianos.
São mostrados os pontos de fixação, exceto para o nervo troclear, que surge da face
dorsal do tronco encefálico.
Existem doze nervos cranianos, pareados bilateralmente, por onde cursam fibras
aferentes e eferentes entre o encéfalo e estruturas periféricas, situadas principalmente
na cabeça e no pescoço. Cada um deles recebe designação específica e são
numerados (em algarismos romanos) de acordo com a seqüência rostro-caudal de sua
fixação encefálica (Fig. 7.1):
I olfativo
II óptico
III oculomotor
IV troclear
V trigêmeo
VI abducente
VII facial
VIII vestibulococlear
IX glossofaríngeo
X vago
XI acessório
XII hipoglosso
71
Os dois primeiros nervos cranianos se prendem diretamente ao telencéfalo, enquanto
os demais se fixam ao tronco encefálico. O sistema olfativo é intimamente associado,
tanto estrutural, como funcionalmente, com as partes do telencéfalo, designadas, em
conjunto, como sistema límbico e, por isso, são discutidos juntos no Cap. 13. O sistema
visual é apresentado no Cap. 12. Os nervos cranianos III e XII estão associados a
diversos núcleos do tronco encefálico, os núcleos dos nervos cranianos, que recebem
aferentes pelos nervos cranianos ou contêm os corpos celulares dos neurônios
eferentes, cujos axônios emergem do encéfalo por nervos cranianos. A localização
desses núcleos é apresentada na Fig. 7.2.
NÚCLEOS DOS NERVOS CRANIANOS AFERENTES
As fibras que conduzem a sensibilidade geral (tato, pressão, dor, temperatura) entram
no tronco encefálico, em sua maioria, pelo nervo trigêmeo, ao nível da ponte, indo
terminar no núcleo sensorial do trigêmeo. Este é um núcleo volumoso, que se estende
por todo o comprimento do tronco encefálico, chegando a penetrar na medula espinhal
cervical. As fibras condutoras das sensibilidades especiais de audição e de
movimento/posição, cursam pelo nervo vestibulococlear. Elas terminam
respectivamente nos núcleos vestibulares e cocleares, localizados no bulbo, na parte
lateral do assoalho do quarto ventrículo ou próximo a ela. Os aferentes viscerais,
incluindo as fibras gustativas, terminam no núcleo solitário, no bulbo.
NÚCLEOS DOS NERVOS CRANIANOS EFERENTES
Com base em sua derivação embriológica, os núcleos dos nervos cranianos eferentes
podem ser divididos em três grupos; cada um deles ocupa uma coluna longitudinal
descontínua.
Núcleos da coluna de células somáticas eferentes
A coluna de células somáticas eferentes fica próxima à linha média, sendo formada
pelos núcleos dos nervos III, IV, VI e XII. O núcleo oculomotor fica no ápice ventral da
substância cinzenta penaquedutal, ao nível do colículo superior. Suas
Fig. 7.2 O tronco encefálico, visto por sua face dorsal.
Essa figura esquemática apresenta as localizações (à esquerda) dos núcleos dos
nervos cranianos aferentes e (à direita) a dos núcleos de nervos cranianos eferentes. A
direita, os núcleos com a mesma cor têm origem embriológica comum. (Modificada,
com autorização, de Barr ML, Kiernan JA 1993 The Human Nervous System, 6. edição
J.B. Lippincott, Filadélfia.)
72
fibras eferentes cursam pelo nervo oculomotor para inervar o músculo elevador da
pálpebra superior, além de todos os músculos extra-oculares, com exceção do oblíquo
superior e do reto lateral, O núcleo troclear também está situado no mesencéfalo, na
borda ventral da substância cinzenta penaquedutal, mas ao nível do colículo inferior.
Suas fibras cursam pelo nervo troclear, para inervar o músculo oblíquo superior do olho.
O núcleo abducente fica localizado na ponte caudal, por baixo do assoalho do quarto
ventrículo. Seus efe- rentes cursam pelo nervo abducente, inervando o músculo reto
lateral. O núcleo do hipoglosso fica no bulbo; suas fibras, por meio do nervo
hipoglosso, inervam os músculos intrínsecos e extrínsecos da língua.
Núcleos da coluna de células braquiomotoras
A coluna celular braquiomotora inerva os músculos estriados derivados dos arcos
branquiais. No tegmento da ponte média fica situado o núcleo motor do trigêmeo, que
fornece fibras para o nervo trigêmeo, inervando os músculos da mastigação e os
músculos tensor do tímpano, tensor do véu do paladar, miloióide e o ventre anterior do
músculo digástrico. No tegmento da ponte caudal, fica o núcleo motor do facial. Este
núcleo inerva os músculos da expressão facial e o músculo estapédio, por meio do
nervo facial. No bulbo, fica o núcleo ambíguo. Este longo núcleo emite fibras motoras,
pelos nervos glossofaríngeo, vago e pela parte craniana do nervo acessório, para
inervar os músculos da faringe e da laringe.
Núcleos das colunas de células parassimpáticas
A coluna de células parassimpáticas é formada por neurônios pré-ganglionares
parassimpáticos, que enviam axônios para os nervos cranianos III, VII, IX e X. O grupo
celular mais rostral é designado como núcleo de Edinger-Westphal, situado na
substância cinzenta periaquedutal do mesencéfalo, adjacente ao núcleo oculomotor.
Seus axônios cursam pelo nervo oculomotor até o gânglio ciliar, onde ficam as células
pós-ganglionares, cujos axônios inervam o esfíncter pupilar e o músculo ciliar do olho.
No tegmento pontino existem dois grupos celulares, os núcleos salivatórios superior e
inferior. O núcleo salivatório superior envia fibras pré-ganglionares para o nervo facial,
que terminam nos gânglios pterigopalatino e submandibular. Fibras pós-ganglionares,
originadas no gânglio pterigo-palatino, inervam as glândulas lacrimais e as membranas
mucosas oral e nasais. As do gânglio submandibular inervam as glândulas
submandibulr e sublingual. O núcleo salivatório inferior envia fibras pré-ganglionares
para o nervo glossofaríngeo. Essas fibras terminam no gânglio ótico, que, por sua vez,
envia axônios pós-ganglionares para a parótida, uma glândula salivar. O maior grupo de
células parassimpáticas pré-ganglionares fica no bulbo, formando o núcleo motor
dorsal do vago. Sua parte rostral fica imediatamente abaixo do assoalho do quarto
ventrículo, lateral ao núcleo do hipoglosso. Suas fibras cursam pelo nervo vago e são
amplamente distribuídas para as vísceras torácicas e abdominais.
NERVOS CRANIANOS
III: NERVO OCULOMOTOR
O nervo oculomotor contém fibras de neurônios motores somáticos, que inervam os
músculos extra-oculares, e de neurônios pré-ganglionares parassimpáticos, que, por
intermédio do gânglio ciliar, controlam o músculo liso, no interior do globo ocular.
Os neurônios motores que inervam os músculos extra- oculares têm seus corpos
celulares no núcleo oculomotor, situado na base da substância cinzenta periaquedutal
do meFig. 7.3 Corte transverso através do mesencéfalo, ao nível do colículo superior. O
esquema mostra a origem e o curso das fibras do nervo oculomotor pelo tronco
encefálico.
73
sencéfalo, ao nível do colículo superior (Fig. 7.3). As fibras dos neurônios préganglionares parassimpáticos emergem do núcleo de Edinger-Westphal vizinho. As
fibras dessas duas origens cursam, ventralmente, pelo tegmento mesencefálico, muitas
delas atravessando o núcleo vermelho, para sair na face medial da cruz do cérebro, na
fossa interpeduncular. O nervo oculomotor passa entre as artérias cerebral posterior e
cerebelar superior, e, em seguida, segue anteriormente, pela parede do seio cavernoso,
antes de ter acesso à órbita, pela fissura orbitária superior, O nervo oculomotor inerva
todos os músculos extra-oculares, com exceção dos músculos oblíquo superior e reto
lateral e, assim, atua para elevar, abaixar e aduzir o globo ocular (Fig. 7.4). Também
inerva o músculo estriado do músculo elevador da pálpebra superior. As fibras dos
neurônios pré-ganglionares parassimpáticos terminam no gânglio ciliar. Daí, fibras pósganglionares cursam pelos nervos ciliares curtos para inervar o esfíncter muscular
pupilar da íris (constrictor) e o músculo ciliar, presente no corpo ciliar.
Fig. 7.4 Movimentos oculares produzidos pelos músculos extra-oculares.
O reflexo pupilar à luz
A quantidade de luz que entra no olho é regulada pelo tamanho da pupila. A iluminação
da retina promove a constricção da pupila, devido à contração do esfíncter muscular
pupilar da íris, reduzindo, desse modo, a quantidade de luz que atinge a retina. Isso é
conhecido como reflexo direto à luz (Fig. 7.5). Mesmo se apenas uma das retinas for
iluminada (como, por exemplo, durante o exame clínico), as pupilas dos dois olhos se
contraem. A constricção da pupila do olho não-iluminado é chamada de reflexo
consensual à luz. O ramo aferente do reflexo à luz é formado por pequeno contingente
de fibras do trato óptico que passam diretamente do olho para a área pré-tectal,
imediatamente rostral ao colículo superior, em vez de se dirigirem para o núcleo
geniculado lateral do tálamo (ver também o Cap. 12). Os neurônios da área pré-tectal
se projetam bilateralmente para o núcleo de Edinger-Westphal, cujas fibras eferentes
cursam pelo nervo oculomotor.
O reflexo de acomodação
A fixação ocular, sobre objeto próximo, por meio da convergência dos eixos ópticos,
envolve a contração concomitante do músculo ciliar, para aumentar a convexidade do
cristalino, o que focaliza a imagem. Também é acompanhada por constricção pupilar.
Esse fenômeno envolve o córtex visual, com fibras córtico-bulbares, ativando
bilateralmente os neurônios parassimpáticos do núcleo de Edinger-Westphal.
Fig. 7.5 Representação esquemática de um corte transverso através da parte mais
rostral do mesencéfalo.
O esquema mostra as vias envolvidas no reflexo pupilar à luz.
74
IV: NERVO TROCLEAR
O nervo troclear só contém fibras de neurônios motores somáticos. Essas fibras
emergem do núcleo troclear, situado na substância cinzenta periaquedutal
mesencefálica, ao nível do colículo inferior (Fig. 7.6). Esses axônios cursam
dorsalmente, em torno da substância cinzenta periaquedutal, cruzando a linha média, O
nervo troclear emerge da face dorsal do tronco encefálico (é o único nervo craniano a
fazê-lo), imediatamente caudal ao colículo inferior. Esse nervo cursa em torno do
pedúnculo cerebral, para atingir a face ventral do encéfalo, passando por entre as
artérias cerebral posterior e cerebelar superior, do mesmo modo como faz o nervo
oculomotor. Em seguida, cursa anteriormente pela parede lateral do seio cavernoso,
entrando na órbita pela fissura orbitária superior. Ele inerva apenas um músculo extraocular, o oblíquo superior, que desloca o globo ocular medialmente para baixo.
Fig. 7.6 Corte transverso através do mesencéfalo, ao nível do colículo inferior.
O esquema mostra a localização do núcleo troclear e o curso das fibras do nervo
troclear.
VI: NERVO ABDUCENTE
O nervo abducente, como o troclear, só contém fibras de neurônios motores somáticos.
Seus corpos celulares de origem ficam situados no núcleo abducente, localizado
abaixo do assoalho do quarto ventrículo, na ponte caudal (Fig. 7.7). Essas fibras
cursam ventralmente pela ponte, emergindo da superfície ventral do tronco encefálico,
na junção entre a ponte e a pirâmide bulbar. Em seguida, o nervo cursa anteriormente,
através do seio cavernoso, entrando na órbita pela fissura orbitária superior, indo
inervar o músculo reto lateral, que produz abdução do globo ocular.
V: NERVO TRIGÊMEO
O nervo trigêmeo tem componentes sensoriais e motores. Representa o principal nervo
sensitivo para a cabeça e, além disso, inerva os músculos da mastigação. Ele se fixa ao
tronco encefálico por meio de duas raízes adjacentes (uma grande, sensorial, e outra
menor, motora), na face ventrolateral da ponte, onde ele se funde com o pedúnculo
cerebelar médio (Fig. 7.1).
As fibras sensoriais do nervo trigêmeo são de neurônios sensoriais primários (primeira
ordem), cujos prolongamentos periféricos são distribuídos, por meio dos ramos
oftálmico, maxilar e mandibular do nervo trigêmeo para diversas estruturas cefálicas
(Fig. 7.8). As sensações de tato, pressão, dor e
Fig. 7.7 Corte transverso através da ponte caudal.
O esquema mostra a localização do núcleo abducente e o curso das fibras do nervo
abducente.
Fig. 7.8 Distribuição superficial das fibras sensitivas nos três ramos do nervo trigêmeo.
75
temperatura são conduzidas da face e do escalpo, da cómea, das cavidades nasais e
oral, inclusive dos dentes e gengivas, e da dura-máter craniana. Além disso, fibras
proprioceptivas, originadas nos músculos da mastigação e da articulação têmporomandibular fazem parte do contingente sensorial desse nervo. Os corpos celulares dos
aferentes do nervo trigêmeo, com exceção dos que conduzem a propriocepção, ficam
localizados no gânglio do trigêmeo (ou semilunar), situado na convergência dos
nervos oftálmico, maxilar e mandibular. Os prolongamentos centrais dessas células
terminam no núcleo sensorial do trigêmeo (Fig. 7.11).
O núcleo sensorial do trigêmeo é uma estrutura volumosa, estendendo-se por todo o
comprimento do tronco encefálico até a parte superior da medula espinhal cervical. É
considerado como formado por três subnúcleos. O núcleo sensorial principal fica no
tegmento pontino, próximo ao ponto de entrada do nervo trigêmeo. O núcleo
mesencefálico se estende rostralmente até o mesencéfalo. O núcleo espinhal (ou
núcleo do trato espinhal do trigêmeo) se estende caudalmente, pelo bulbo, indo até a
medula, onde se torna contínuo com substância gelatinosa, da qual é considerado ser o
homólogo no tronco encefálico.
Lesões dos nervos cranianos III, IV e VI
A paralisia do nervo craniano III (Fig. 7.9), decorrente de lesão do núcleo oculomotor,
no mesencéfalo, ou de compressão desse nervo, por aneurisma ou tumor, em seu
curso perifénco, é caracterizada por queda da pálpebra (ptose), dilatação da pupila, que
não responde à luz e à acomodação, além de incapacidade de mover o globo ocular
para cima, para baixo e em direção medial (adução).
A paralisia do nervo craniano VI (Fig. 7.10), decorrente de lesão do núcleo abducente,
na ponte, ou de compressão do nervo, em seu percurso periférico, por aneurisma ou
tumor, produz incapacidade de movimentar o globo ocular na direção lateral (abdução).
Paralisias combinadas do terceiro, quarto e sexto nervos cranianos ocorrem como
conseqüência de lesões ao longo de seus percursos periféricos, onde esses nervos
cursam adjacentes entre si, como dentro do seio cavernoso, na entrada da órbita
(fissura orbitária superior) e dentro da órbita. Aqui, eles são vulneráveis à compressão
por tumores e aneurismas. Os efeitos dessas lesões são os seguintes:
• ptose
• dilatação da pupila, que deixa de responder à luz ou apresentar acomodação
• paralisia de todos os movimentos oculares.
Fig. 7.9 Paralisia do nervo oculomotor (III) direito.
A parte superior da figura mostra ptose à direita. A parte inferior mostra que, com
elevação da pálpebra, o globo ocular pode ser visto abduzido e a pupila dilatada.
Fig. 7.10 Paralisia do nervo abducente (VI) esquerdo.
Ao olhar na direção da seta, o olho esquerdo deixa de abduzir.
76
Fig. 7.11 Esquema do tronco encefálico, indicando a localização do núcleo
sensorial do trigêmeo e suas principais conexões.
(Modificado, com permissão, de Barr ML, Kiernan JA 1993 The Human Nervous System
6. edição, JB Lippincott, Filadélfia.)
Existe distribuição segregada da terminação das fibras aferentes nos núcleos
trigeminais. As fibras condutoras de tato e pressão terminam no núcleo principal. As
que conduzem dor e temperatura terminam no núcleo espinhal, atingindo seu término
ao descerem pelo trato espinhal do trigêmeo, situado imediatamente superficial ao
núcleo. Na medula espinhal cervical, o trato espinhal do trigêmeo se toma contínuo com
o trato de Lissauer, que contém aferentes funcionalmente homólogos, originados de
nervos espinhais, antes de terminarem na ponta dorsal. Os aferentes proprioceptivos
dos músculos da mastigação e da articulação têmporo-mandibular não têm seus corpos
celulares de origem no gânglio trigeminal, como seria de se esperar, mas, sim, no
núcleo mesencefálico do trigêmeo. São os únicos aferentes primários a ter seus corpos
celulares no interior do sistema nervoso central. Axônios emergentes de neurônios de
segunda ordem no núcleo trigeminal decussam para formar o trato trigêmino-talâmico
(lemnisco trigeminal) contralateral. Este trato termina no núcleo ventral posterior
contralateral do tálamo que, por sua vez, envia fibras para o córtex sensorial do lobo
parietal. Além disso, o núcleo do trigêmeo envia fibras para o cerebelo e forma
conexões reflexas com determinados grupos celulares motores do tronco encefálico.
Proeminente entre esses, é o núcleo facial, por meio do qual são mediados os trejeitos
faciais e fechamento das pálpebras (reflexo comeano), em resposta à estimulação
nóxia no território inervado pelo trigêmeo.
Os axônios motores do nervo trigêmeo emergem de células do núcleo motor do
trigêmeo, situado no tegmento pontino, medial ao núcleo sensorial principal. Esses
axônios emergem da ponte pela raiz motora do trigêmeo e, em seguida, juntam- se ao
ramo mandibular. Eles inervam diversos músculos; os mais importantes são os
músculos da mastigação (masseter e temporal, que elevam a mandíbula, e os
pterigóides lateral e medial, que a abaixam).
Lesões do nervo trigêmeo
A infecção por herpes zoster das raízes sensoriais do nervo trigêmeo (zona [shingles])
provoca dor e erupção de vesículas localizadas no dermátomo inervado por um ou mais
dos ramos (oftálmico, maxilar, mandibular) do nervo trigêmeo.
Na siringobulbia, a cavitação central do bulbo, caudal ao quarto ventrículo, produz
destruição compressiva preferencial das fibras trigêmino-talâmicas que decussam,
causando perda seletiva da sensibilidade à dor e à temperatura (perda sensorial
dissociada) na face (confrontar com siringomielia).
VII: NERVO FACIAL
O nervo facial tem componentes sensitivos, motores e parassimpáticos (Fig. 7.12). Ele
chega ao tronco encefálico pela face
77
Fig. 7.12 As fibras componentes do nervo facial e sua distribuição periférica.
(vermelho = motoras, verde = sensoriais, azul = parassimpáticas)
ventrolateral da ponte caudal, próximo da junção ponto-bulbar, na região conhecida
como ângulo cerebelo-pontino. Esse nervo é formado por duas raízes, a mais lateral
(chamada, às vezes, de nervo intermédio) contém fibras sensoriais e parassimpáticas,
enquanto a mais medial só contém fibras motoras.
As fibras sensoriais do nervo facial conduzem a sensibilidade gustativa dos dois terços
anteriores da língua, do assoalho da boca e do palato, além da sensibilidade cutânea
de parte do ouvido externo. Os corpos celulares dos neurônios aferentes primários
ficam no gânglio geniculado, dentro do canal facial da parte petrosa do osso temporal.
Os prolongamentos centrais das fibras gustativas terminam na parte rostral do núcleo
solitário, no bulbo. As fibras ascendentes, que partem do núcleo solitário, se projetam
para o núcleo ventral posterior do tálamo que, por sua vez, envia fibras para o córtex
sensorial do lobo parietal. As fibras condutoras da sensibilidade cutânea terminam no
núcleo trigeminal.
As fibras motoras do nervo facial têm origem no núcleo motor do facial, no tegmento
pontino caudal (Fig. 7.13). Esses axônios passam, inicialmente, em sentido dorsal,
seguindo por sobre o núcleo abducente, por baixo do assoalho do quarto ventrículo,
antes de emergirem do tronco encefálico pela raiz motora do nervo facial. Essas fibras
motoras são distribuídas para os músculos da expressão facial, e para os músculos
platisma, estiloióideo, para o ventre posterior do múculo digástrico e para o músculo
estapédio do ouvido médio.
O núcleo motor do facial recebe aferentes de outras áreas do tronco encefálico, para
mediação de determinados reflexos, bem como do córtex cerebral. São formadas
conexões reflexas que medeiam o fechamento palpebral em resposta a estímulos
visuais ou a estimulação táctil da córnea (reflexo corneano), por fibras oriundas,
respectivamente, do colículo superior e do núcleo sensorial do trigêmeo. Além dessas,
fibras oriundas do núcleo olivar superior, parte da via auditiva central, promovem a
contração reflexa do estapédio, em resposta a sons intensos.
Fig. 7.13 Corte transverso através da ponte.
O esquema mostra a origem e o curso das fibras motoras do nervo facial.
78
Fibras córtico-bulbares, originadas de áreas corticais motoras, inervam o núcleo motor
do facial. As que controlam os neurônios motores que inervam os músculos da parte
superior da face (frontal, orbicular do olho) têm distribuição bilateral. As que controlam
os neurônios motores que inervam os músculos da parte inferior da face são totalmente
cruzadas. Por conseguinte, as lesões unilaterais do neurônio motor superior produzem
paralisia dos músculos inferiores da face.
As fibras pré-ganglionares parassimpáticas têm origem no núcleo salivatório superior,
na ponte. Essas fibras saem do tronco encefálico pela raiz sensorial do nervo facial
(nervo intermédio). Daí, elas passam para gânglios parassimpáticos — os gânglios
submandibular e pterigo-palatino — onde fazem sinapses com neurônios pósganglionares. As do gânglio submandibular inervam as glândulas salivares
submandibular e sublingual. As fibras pós-ganglionares do gânglio pterigopalatino
inervam a glândula lacrimal e as membranas mucosas nasais e oral.
Paralisia de BeIl
A paralisia de BeIl representa uma lesão inflamatória unilateral aguda do nervo facial,
através de seu curso pelo crânio. E sentida dor em tomo da orelha e existe paralisia
unilateral dos músculos faciais, com incapacidade de fechar as pálpebras, ausência do
reflexo comeano, hiperacusia no lado afetado e perda da sensibilidade gustativa nos
dois terços anteriores da língua. Quando o vírus herpes zoster é o agente inflamatório,
fica aparente uma erupção vesicular no canal auditivo externo e nas membranas
mucosas da orofaringe (síndrome de Ramsay Hunt).
VIII: NERVO VESTIBULOCOCLEAR
O nervo vestibulococlear é um nervo sensitivo que conduz impulsos a partir do ouvido
interno. Ele tem dois componentes: o nervo vestibular, condutor de informação
relacionada à posição e à movimentação da cabeça e o nervo coclear, condutor de
informação auditiva. Esses dois componentes contêm os axônios de neurônios
sensitivos de primeira ordem, cujos dendritos fazem contato com células ciliadas no
aparelho vestibular ou no aparelho auditivo do ouvido interno. Esses dois componentes
passam, juntos, pelo meato auditivo interno (por onde também passa o nervo facial) e
se prendem ao tronco encefálico na junção entre o bulbo e a ponte, na região
conhecida como ângulo cerebelo-pontino.
O nervo vestibular
O nervo vestibular é formado por fibras cujos dendritos contactam células chiadas das
regiões vestibulares do labirinto membranoso e seus corpos celulares situados no
gânglio vestibular, dentro do meato auditivo interno. Os prolongamentos centrais das
fibras vestibulares terminam, em sua maioria, nos núcleos vestibulares, no bulbo rostral.
Esses núcleos são em número de quatro (núcleos vestibulares superior, inferior, medial
e lateral), situados bastante próximos entre si, por baixo da parte lateral do assoalho do
quarto ventrículo.
Os núcleos vestibulares estabelecem contato com diversas outras regiões, para o
controle da postura, manutenção do equilíbrio, coordenação dos movimentos da cabeça
e dos olhos e conhecimento consciente da estimulação vestibular. As fibras do núcleo
vestibular lateral (núcleo de Deiters’) descem ipsilateralmente pelo trato vestíbuloespinhal lateral. Os núcleos vestibulares também contribuem com fibras para o
fascículo longitudinal medial (FLM), que cursa por todo o tronco encefálico, atingindo
a medula espinhal. O componente descendente do FLM também é designado como
trato vestíbulo-espinhal medial. As fibras vestíbulo-espinhais influenciam a atividade
dos neurônios motores espinhais, relacionados com o controle da postura do corpo e do
equilíbrio. A parte ascendente do fascículo longitudinal medial estabelece conexões
com os núcleos dos nervos abducente, troclear e oculomotor, para a coordenação dos
movimentos da cabeça e dos olhos. Algumas fibras eferentes dos núcleos vestibulares
passam pelo pedúnculo cerebelar inferior, até o lobo flóculo-nodular do cerebelo, que é
relacionado ao controle do equilíbrio. Outras fibras ascendem para o tálamo
contralateral (núcleo ventral posterior) que, por sua vez, projeta-se para o córtex
cerebral. A região cortical, responsável pelo conhecimento consciente da sensibilidade
vestibular ainda não está definida, mas provavelmente fica próxima da área da
“cabeça”. no córtex sensorial do lobo parietal, ou adjacente ao córtex auditivo, no lobo
temporal.
O nervo coclear
O nervo coclear é formado por fibras cujos dendritos contactam células ciliadas do
órgão de Corti, no interior do ducto coclear do ouvido interno. Os corpos celulares
dessas células ficam situados no interior da cóclea e, em seu conjunto, formam o
gânglio espiral. O nervo coclear junta-se ao tronco encefálico ao nível do bulbo rostral.
Suas fibras se bifurcam, terminando nos núcleos cocleares dorsal e ventral, situados
próximos ao pedúnculo cerebelar inferior. Desse ponto, a via auditiva ascendente para
o tálamo e para o córtex cerebral (Fig. 7.14) é bastante mais complicada e variável que
a via para as sensibilidades gerais.
Existem diversas localizações, entre o bulbo e o tálamo, onde os axônios podem fazer
sinapses, e nem todas as fibras dessa via seguem um padrão geral. A partir dos
núcleos cocleares, fibras de neurônios de segunda ordem ascendem até a ponte,
algumas cruzando para o tegmento pontino do lado oposto, formando o corpo
trapezóide. Nesse nível, algumas fibras terminam no núcleo olivar superior. Esse
núcleo é o local de origem das fibras olivococleares, que saem do tronco encefálico
pelo nervo vestibulococlear, terminando no órgão de Corti. Elas têm função inibitória,
atuando para modular a transmissão de informação auditiva para o nervo coclear.
Partindo do núcleo olivar superior, fibras ascendentes formam o lemnisco lateral, que
cursa pelo tegmento pontino, para terminar no colículo inferior do mesencéfalo.
Algumas fibras do lemnisco lateral terminam em um pequeno núcleo pontino, chamado
de núcleo do lemnisco lateral. O núcleo olivar superior e o núcleo do lemnisco lateral
são considerados como estabelecendo conexões reflexas com neurônios motores dos
núcleos motores do trigêmeo e do facial, mediando a contração dos músculos tensor do
tímpano e estapé79
Fig. 7.14 – Principais conexões ascendentes do componente auditivo do nervo
vestibulococlear.
80
dio, em resposta a sons muito intensos, O colículo inferior, por sua vez, envia axônios
para o núcleo geniculado medial do tálamo. A etapa final da via auditiva ascendente é
formada por axônios que se originam no núcleo geniculado medial, passando pela
cápsula interna, para atingir o córtex auditivo primário, no lobo temporal. Esse córtex
fica localizado, em sua maior parte, nos giros de Heschl, situados na superfície dorsal
do giro temporal superior e, por isso, ocultos, em grande parte, dentro da fissura
lateral. Por toda a projeção auditiva ascendente, existe uma representação chamada
“tonotópica” da cóclea, que é análoga à organização “somatotópica” das vias para a
sensibilidade geral. No tronco cerebral, algumas fibras ascendentes decussam,
enquanto outras não o fazem. A representação da cóclea é, assim, essencialmente
bilateral em todos os níveis rostrais aos núcleos cocleares. A região do lobo temporal,
circundante do córtex auditivo primário, é conhecida como córtex auditivo de
associação ou área de Wernicke. E aí que a informação auditiva é interpretada,
recebendo significação contextual. A área de Wernicke é importante para o
processamento da linguagem pelo cérebro (ver Cap. 10).
IX: NERVO GLOSSOFARÍNGEO
O nervo glossofaríngeo é sobretudo um nervo sensitivo, embora também contenha
fibras pré-ganglionares parassimpáticas e algumas fibras motoras. Ele se prende ao
tronco encefálico como seqüência linear de pequenas radículas, laterais à oliva, no
bulbo rostral.
As fibras aferentes do nervo glossofaríngeo conduzem informação oriunda de:
• receptores para a sensibilidade geral na faringe, no terço posterior da língua, na tuba
de Eustáquio e no ouvido médio;
• papilas gustativas na faringe e no terço posterior da língua;
• quimiorreceptores no corpúsculo carotídeo e de barorreceptores no seio carotídeo.
No tronco cerebral, as fibras aferentes para a sensibilidade geral terminam no núcleo
sensorial do trigêmeo. As fibras condutoras da sensibilidade táctil, originadas na faringe
e parte posterior da língua, são importantes na mediação do reflexo do engasgo, por
meio de conexões com o núcleo ambíguo e com o núcleo hipoglosso. As fibras
viscerais e gustativas do nervo glossofaríngeo terminam no núcleo solitário, no bulbo.
O componente motor do nervo glossofaríngeo é muito pequeno. Origina-se de células
na parte rostral do núcleo ambíguo, no bulbo, e inerva apenas um músculo, o
estilofaríngeo, que participa da deglutição.
As fibras pré-ganglionares parassimpáticas, no nervo glossofaríngeo, originam-se no
núcleo salivatório inferior, no bulbo rostral. Elas fazem sinapses com neurônios pósganglionares no gânglio ótico que, por sua vez, inervam a glândula salivar parótida.
Neuroma acústico
Um neuroma acústico é um tumor benigno do oitavo nervo craniano, que causa
compressão desse nervo e das estruturas adjacentes no ângulo cerebelo-pontino.
Ocorrem ataques de tonteiras, acompanhados por surdez profunda, e, com o
crescimento do tumor, ataxia e paralisia de nervos cranianos (especialmente dos V-VII)
e dos membros. Neuromas acústicos uni- e bilaterais ocorrem na doença hereditária
neurofibromatose, na qual tumores dos nervos periféricos e da pele (neurilemoma e
neurofibroma) podem causar defeitos cutâneos e deformidades.
X: NERVO VAGO
As radículas do nervo vago se prendem à face lateral do bulbo, imediatamente caudais
às do nervo glossofaríngeo. O nervo vago contém fibras aferentes, motoras e
parassimpáticas.
As fibras aferentes do vago conduzem informação originada de:
• receptores da sensibilidade geral na: faringe, laringe, esôfago, membrana timpânica,
meato auditivo externo e parte da concha do ouvido externo;
• quimiorreceptores dos corpúsculos aórticos e barorreceptores do arco aórtico;
• receptores amplamente distribuídos nas vísceras torácicas e abdominais.
No tronco encefálico, os receptores para a sensibilidade geral terminam no núcleo
sensorial do trigêmeo, enquanto os aferentes viscerais terminam no núcleo solitário.
As fibras motoras do vago (Fig. 7.15) se originam no núcleo ambíguo, no bulbo. Elas
inervam os músculos do palato mole, da faringe, da laringe e da parte superior do
esôfago. O núcleo ambíguo é, assim, de importância crucial para o controle da
deglutição e da fala. Por convenção, os eferentes mais caudais, que emergem do
núcleo ambíguo, são considerados como saindo do tronco encefálico pelas raízes
cranianas do nervo acessório, mas elas passam para o próprio nervo vago, no nível do
forâmen jugular (Fig. 7.15).
As fibras parassimpáticas do nervo vago se originam do núcleo motor dorsal do vago,
situado no bulbo, imediatamente abaixo do assoalho do quarto ventrículo. Elas são
distribuídas amplamente por todos os sistemas cardiovascular, respiratório e
gastrointestinal.
XI: NERVO ACESSÓRIO
O nervo acessório tem função exclusivamente motora. Ele é composto por duas partes:
a craniana e a espinhal. A parte craniana emerge da face lateral do bulbo, como uma
série linear de radículas, situadas imediatamente caudais às radículas do nervo vago
(Fig. 7.15). A raiz craniana do nervo acessório contém fibras que se originam da parte
caudal do núcleo ambíguo, no bulbo. No nível do forâmen jugular, essas fibras entram
no nervo vago, sendo distribuídas para os músculos do palato mole, faringe e laringe.
A raiz espinhal do nervo acessório se origina em neurônios motores situados na ponta
ventral da substância cinzenta medular dos níveis Cl a C5 (Fig. 7.15). Esses axônios
saem da medula, não pelas raízes ventrais de nervos espinhais, mas, sim, por meio de
uma série de radículas que emergem da face lateral da medula, a meio caminho entre
as raízes dorsais e ventrais. Essas radículas cursam rostralmente, coalescendo à me81
Fig. 7.15 Esquema do bulbo caudal e da medula espinhal rostral.
A figura mostra a origem e o curso das fibras motoras dos nervos vago e acessório.
dida que ascendem, até entrar na cavidade craniana pelo forâmen magno. Ao lado do
bulbo, a raiz espinhal do nervo acessório se junta, por pequena extensão, à raiz
craniana, mas as fibras componentes logo se separam, quando o nervo sai da cavidade
craniana pelo forâmenjugular. Nesse ponto, as fibras da raiz craniana, derivadas do
núcleo ambíguo, juntam-se às do vago, para serem distribuídas junto com elas. As
fibras da raiz espinhal passam para os músculos estemomastóideo e trapézio, que
atuam nos movimentos da cabeça e dos ombros.
XII: NERVO HIPOGLOSSO
O nervo hipoglosso também tem função exclusivamente motora. Ele inerva os músculos
extrínsecos e intrínsecos da língua e, desse modo, provoca o movimento e a mudança
da posição da língua. Seus axônios se originam no núcleo hipoglosso, situado
imediatamente abaixo de assoalho do quarto ventrículo, próximo à linha média (Fig.
7.16). Esses axônios cursam medialmente pelo bulbo, emergindo na sua face
ventrolateral como uma série linear de radículas, localizada entre a pirâmide e a oliva, O
núcleo hipoglosso recebe aferentes do núcleo solitário e do núcleo sensorial do
trigêmeo; estes núcleos participam do controle dos movimentos reflexos da mastigação,
sucção e deglutição. Ele também recebe fibras córtico-bulbares do córtex motor
contralateral, que promovem os movimentos voluntários da língua, como os que ocoem
na fala.
82
Doença do neurônio motor e lesões dos nervos IX-XlI
Na doença do neurônio motor, um distúrbio degenerativo crônico em indivíduos com
mais de cinqüenta anos, os tratos córtico-bulbares, que se projetam para os núcleos
ambíguo e hipoglosso, degeneram, levando à disfonia (dificuldade na fonação),
disfagi.a (dificuldade na deglutição), disartria (dificuldade na articulação das palavras),
fraqueza e espasticidade da língua (paralisia pseudobulbar). Também ocorre
degeneração dos próprios núcleos ambíguo e hipoglosso, causando disfonia, disfagia,
disartr.ia, além de fraqueza, atrofia e fasciculação da língua (paralisia bulbar).
O nono, décimo, undécimo e duodécimo nervos podem ser lesados por compressão,
em seu percurso periférico, ao saírem do crânio, passando pelos forâmens da base
craniana. Os tumores, nessas áreas, causam disfonia, fraqueza unilateral, atrofia e
fasciculações da língua, com o reflexo do engasgo ficando deprimido, junto com atrofia
unilateral dos músculos esternomastóideo e trapézio
Fig. 7.16 Corte transverso através do bulbo.
A figura mostra a origem e o curso das fibras do nervo hipoglosso.
84