Sistema imune inato
Tecnologias de Informação e Comunicação na Educação
Professora Ana Paula Peconick
Tutor Karlos Henrique Martins Kalks
Lavras/MG
2011
1|Página
Ficha catalográfica preparada pela Divisão de Processos
Técnicos da Biblioteca Central da UFLA
Espaço a ser preenchido pela biblioteca
[A ser preenchido posteriormente]
Espaço a ser preenchido pelo CEAD
______________Sistema imune inato______________
Índice
UNIDADE 3 ............................................................................ 6
3.1 Introdução....................................................................... 7
3.2 Imunidade inata .............................................................. 8
3.2.1 Padrões de reconhecimento na resposta imune inata10
3.2.2 Receptores solúveis ................................................. 12
3.2.2.1 As colectinas ..................................................... 12
3.2.2.2 Cascata complemento ........................................ 13
3.2.2.3 Pentraxinas ....................................................... 15
3.2.2.4 Proteínas ligadas ao lipopolissacarídeos e CD14 15
3.2.3 Receptores associados a células .............................. 15
3.2.3.1 Receptores Toll-like ........................................... 15
3.2.3.3 Receptores de peptídeos formil.......................... 19
3.2.4 Receptores intracelulares ........................................ 19
3.3 Citocinas e quimiocinas.................................................. 20
3.4 Células do sistema inume inato ...................................... 22
3.4.1 Mucosa epitelial – primeira linha de defesa .............. 22
3.4.2 Células fagocitárias: neutrófilos e macrófagos ......... 24
3.4.3 Células dendríticas (DCs) ......................................... 26
3.4.4 Células natural killers (NK) ...................................... 28
3.4.5 Mastócitos ............................................................... 30
3.5 Conclusão ...................................................................... 31
3.6 Bibliografia .................................................................... 32
4|Página
______________Sistema imune inato______________
5|Página
______________Sistema imune inato______________
UNIDADE 3
OBJETIVO: Conhecer o sistema imunológico inato, destacando seus
componentes, funções e interações.
6|Página
______________Sistema imune inato______________
3.1 Introdução
Imunidade refere-se à habilidade global de um
hospedeiro resistir à ação predatória de micróbios que
podem
prejudicá-lo.
A
própria
definição
da
palavra
patógeno, organismo que pode superar as defesas do
corpo
e
induzir
alterações
deletérias,
demonstra
a
importância da existência de mecanismos eficazes de
proteção do hospedeiro. Dois sistemas constituem a
defesa do corpo contra a ação de patógenos. Eles podem
ser divididos por uma linha evolucionária germinal antiga,
na
qual
um,
o
mais
antigo
desenvolveu-se
para
proporcionar uma resposta rápida (inespecífica) à invasão
pelos microrganismos. Já o outro, de evolução mais
tardia, responde mais lentamente a infecção, porém de
forma específica. Esses dois sistemas imunes que dão
proteção ao organismo são conhecidos como imunidade
inata
e
imunidade
adaptativa
(adquirida),
respectivamente.
A imunidade inata surgiu com os primeiros seres
multicelulares, sendo que o benefício da sua existência é
destacado pelo fato da maioria dos organismos, sobre
nosso planeta, sobreviver apenas com este tipo de
imunidade, como por exemplo, as plantas e os insetos. Já
a
imunidade
mandibulados,
adaptativa
e
possui
surgiu
como
com
os
principal
vertebrados
vantagem
reconhecer praticamente qualquer antígeno existente,
além da geração de memória imunológica, diferentemente
do sistema inato, que reconhece apenas um número
limitado de antígenos.
Apesar desta divisão entre os tipos de imunidade,
ocorre um contínuo entre estes dois sistemas. Assim
7|Página
______________Sistema imune inato______________
sendo, existem complexos mecanismos de interação entre
a imunidade inata e a adaptativa. O que faz com que as
duas trabalhem em conjunto para a defesa do organismo.
3.2 Imunidade inata
A imunidade inata é primeira linha de defesa do
hospedeiro
contra
patógenos.
Ela
é
constituída
principalmente por elementos mecânicos, químicos e
celulares. A epiderme e as mucosas estão entre os
elementos mecânicos que funcionam como barreiras para
a
entrada
do
patógeno.
Associadas
a
elas
estão
propriedades fisiológicas como baixo pH do estômago, a
ação ciliar, motilidade, descamação e a secreção de
muco, produção da saliva e da lágrima que contém
lisozima. Elementos químicos podem ser divididos em três
subcomponentes:
moléculas
de
reconhecimento
de
padrões, as quais podem ser solúveis ou associadas às
células; proteínas ou peptídeos que hidrolisam micróbios;
e
citocinas
e
quimiocinas
que
regulam
a
resposta
imunológica. Já o elemento celular é constituído por
células epiteliais, mastócitos, células dendríticas, células
fagocitárias como os macrófagos e granulóctios, células
Natural killers e células Tγδ.
A imunidade inata é responsável pela construção da
resposta inflamatória. Esta é ocasionada em uma primeira
instância por macrófagos, leucócitos polimorfonucleares e
mastócitos
relacionados
através
à
de
resposta
receptores
imune
nestas
inata.
O
células
agente
inflamatório age sobre os tecidos e induz a liberação de
mediadores químicos que sobre tais receptores, produzem
aumento da permeabilidade vascular e exsudação de
8|Página
______________Sistema imune inato______________
plasma e células sanguíneas para o interstício. Dessa
forma este mecanismo possibilita a defesa contra muitos
agentes agressores. Contudo, a reação inflamatória pode
em alguns casos levar a processos fisiológicos que
causam dano ao tecido do hospedeiro.
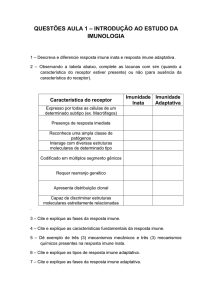
Utilizando várias fontes de pesquisa, escreva as principais
diferenças entre sistema imune inato e sistema imune
adaptativo na tabela abaixo. Compare sua pesquisa com a de
outros colegas.
Tabela 1.
Principais
diferenças
entre
imunidade
inata e
adquirida.
Imunidade inata
Imunidade adquirida
9|Página
______________Sistema imune inato______________
3.2.1 Padrões de reconhecimento na resposta
imune inata
Após
causando
o
patógeno
danos,
aderir
sinais
são
à
superfície
gerados
epitelial
ocasionando
a
produção de quimiocinas, citocinas, prostaglandinas e
leucotrienos pelo epitélio. Estas substâncias indicam dano
no tecido. Células dendríticas e macrófagos também
interagem com o microrganismo invasor, através da
detecção do patógeno por moléculas nestas células
especializadas no reconhecimento de estruturas padrões
presentes
nos
patógenos.
Este
mecanismo
de
reconhecimento é a base da ativação do sistema imune
inato.
Patógenos
são
caracterizados
por
arranjos
específicos de moléculas chaves chamados de padrões
moleculares associados aos patógenos (PAMPs) que são
reconhecidos pelos receptores de reconhecimento de
padrões (PRRs). Diferentes PRRs reagem com específicos
PAMPs mostrando distintos padrões de expressão e vias
de sinalização ativas específicas, o que gera distintas
respostas a diferentes patógenos.
Os
PAMPs
são
estruturas
vitais
das
células
microbianas, as quais têm alterado pouco durante a
evolução. Assim, representam os principais alvos no
reconhecimento do sistema imune inato. Dentre estas
estruturas destacam-se os lipopolissacarídeos e ácidos
teicólicos que estão presentes nas células bacterianas
gram-negativas e gram-positivas, respectivamente; os
motivos não metilados CpGs, os quais caracterizam DNAs
de células bacterianas; RNA de fita-dupla representando a
10 | P á g i n a
______________Sistema imune inato______________
assinatura estrutural de vírus de RNA; e os polímeros de
manose que são componentes da parede celular de
células de leveduras. Nenhuma dessas estruturas são
sintetizadas pelo organismo hospedeiro e todas elas são
compartilhadas por grandes grupos de patógenos e são
essenciais para a sua fisiologia.
Os PRRs são encontrados sobre muitas das células
do
sistema
imune
inato
incluindo
células
epiteliais
macrófagos, monócitos, granulócitos, mastócitos e células
dendríticas. Eles podem ser expressos sobre a superfície
das
células,
em
compartimentos
intracelulares,
ou
secretados na circulação sanguínea e fluídos teciduais. As
principais
funções
ativação
do
dos
sistema
PRRs
incluem
complemento
e
opsonização,
casacata
de
coagulação, fagocitose, ativação de vias sinalizatórias da
inflamação e indução da apoptose. Existem diferentes
famílias
de
PRRs
como
os
receptores
scavenger,
receptores Toll-like (TLR), receptores de peptídeos formil,
receptores
de
manose
e
glicanos,
receptores
do
complemento (CR3) e CD14.
Outra maneira do sistema imune inato perceber a
presença de patógenos é detectar os DAMPs (padrões
moleculares
associados
ao
perigo),
os
quais
são
metabólicos comuns em conseqüência da inflamação e
infecção. A figura 1 esquematiza as principais formas de
detecção de PAMPs pelo sistema imune inato e suas
conseqüências.
11 | P á g i n a
______________Sistema imune inato______________
Figura 1. Receptores padrões de reconhecimento e seus papéis
na resposta imune inata e adaptativa (modificado de BASSET
et al., 2003).
3.2.2 Receptores solúveis
3.2.2.1 As colectinas
A família das colectinas inclui proteínas surfactantes
A, D (SP-A, SP-D) e lecitina ligada a manose (MBL). Estes
receptores se ligam ao glicoconjugados bacterianos e
atuam como opsoninas. SP-A e SP-D, primeiramente,
foram relacionadas a homeostase lipídica, mas agora
sabe-se que elas interagem com um grande número de
estruturas bacterianas e fúngicas.
As colectinas são caracterizadas pela presença de
uma região colágena e um domínio lecitina tipo C (CTL).
Tipicamente elas formam receptores oligoméricos.
12 | P á g i n a
______________Sistema imune inato______________
3.2.2.2 Cascata complemento
As
proteínas
associadas
a
MBL,
ligam-se
a
polímeros de manose das paredes celulares microbianas e
são membros da família das colectinas. As lecitinas têm
um importante papel na ativação da cascata complemento
e elas determinam uma das três vias de ativação do
complemento, chamada via das lecitinas. As outras duas
vias são: via clássica, ativada por complexos antígenoanticorpo e a via alternativa, desencadeada pela ativação
do composto C3 na superfície de patógenos, esta última
via também importante como mecanismo imediato de
defesa do organismo contra agentes infecciosos, logo
fazendo parte da resposta imune inata.
Quando ligadas a resíduos de manose, MBL sofre
uma alteração conformacional que resulta na clivagem
dos
compostos
C4
e
C2
para
formar
o
complexo
convertase C4b2a. Por ação deste o C3 é clivado em dois
fragmentos C3a e C3b. C3b se liga ao C5 fixando-o junto
a membrana do agente agressor. Então uma convertase
chamada C3/C5 cliva o C5 em dois novos fragmentos
(C5a e C5b). C5b possui dois sítios de ligação um que liga
ao C6 e outro que se liga a membrana. O complexo C5b6,
liga-se ao C7 gerando C5b67 que se liga ao C8, formando
um complexo que induz a polimerização do C9. Assim,
este último forma um complexo polimérico de 12 a 16
unidades que constituem canais anfipáticos, hidrofóbicos
externamente
e
hidrofílicos
conhecido como complexo de
internamente,
ataque
também
a membrana,
reponsável pela morte do patógeno sobre o qual ele foi
ativado.
13 | P á g i n a
______________Sistema imune inato______________
Na
via
alternativa
de
ativação
do
sistema
complemento o composto C3b é o agente desencadeador
da cascata. Este composto pode ser gerado de maneira
espotânea porém em baixas proporções. Quando ligado a
superfície do patógeno ele se mantém estável, dessa
forma, possibilita a ligação ao fator B pela ação catalítica
do fator D, produzindo assim o complexo C3bBb, outro
tipo de convertase sobre a superfície do patógeno. O
C3bBb clica o C3 em C3a e C3b, que pode continuar a
cascata de reações com C5, C6, C7, C8 e C9 como
descrito no parágrafo acima. Em termos da “limpeza” do
organismo, a presença de C3b sobre a superfície do
patógeno
pode
fazê-lo
atuar
como
uma
opsonina,
aumentando a absorção pelo fagócitos através de seus
receptores de C3b.
A via clássica de ativação do sistema complemento
requer
a
presença
caracterizando-a
de
como
anticorpos
parte
do
para
ativação,
sistema
imune
adaptativo. Contundo, ela também pode ser ativada pela
proteína reativa C (CRP) e assim também ser considerada
como parte da resposta imune inata. Existem também os
anticorpos
naturais
que
são
IgM
secretadas
espontaneamente e que com freqüência reagem com
polissacarídeos microbianos. Estes anticorpos também
podem ser caracterizados como componentes da resposta
imune inata.
Revise o sistema complemento. Procure identificar seus
componentes e a forma de atuação de cada um deles.
14 | P á g i n a
______________Sistema imune inato______________
3.2.2.3 Pentraxinas
A característica básica desta família é a estrutura
pentamérica cíclica de suas moléculas. Ela inclui proteínas
curtas como a proteína reativa C e soro amilóide P (SAP)
e
grandes
pentraxinas.
CRP
e
SAP
atuam
como
opsoninas, além de poderem ativar o componente C1q da
via clássica do sistema complemento por se ligarem a ele.
Longas pentraxinas são produzidas por uma variedade de
células imunológicas e epiteiais.
3.2.2.4
Proteínas
ligadas
ao
lipopolissacarídeos e CD14
As proteínas ligadas a lipopolissacarídeos (LBP)
estão dispersas no soro e rapidamente se ligam aos LPS,
facilitando a transferência deles para o receptor CD14
sobre os macrófagos. Contudo, este receptor não possui
um domínio intracitoplasmático, sendo necessário então a
associação a uma
receptor
4)
o
outro receptor
qual
é
responsável
o TLR4 (Toll-like
pela
sinalização
intracelular que levará a secreção de citocinas pelos
macrófagos.
3.2.3 Receptores associados a células
3.2.3.1 Receptores Toll-like
Durante os anos noventa foi desmonstrado que a
proteína Toll em espécies de Drosophila está envolvida na
15 | P á g i n a
______________Sistema imune inato______________
defesa destes insetos contra infecções causadas por
fungos. Desde então proteínas similares foram descritas
em
mamíferos.
Estas
diferem
de
cada
outra
na
especificidade dos ligantes, padrões de expressão, e nos
genes alvos que podem induzir. Muitas TLRs estão
envolvidas no reconhecimento de uma variedade de
PAMPs.
Receptores
diméricas
Toll-like
existem
(heterodímeros
ectodomínios
de
TLRs
ou
são
como
proteínas
homodímeros).
compostos
de
Os
motivos
repetidos ricos em leucina, enquanto o componente
citosólico, chamado Toll/IL-1 receptor-like domain (TIR),
está envolvido na sinalização. Após a ligação de produtos
microbianos
com
os
TLRs
ocorre
a
sinalizações
metabólicas como a ativação de fator nuclear κβ, ativação
da proteína 1, fator 3 regulador de interferon, e outros
fatores transcripcionais. Dessa forma, ocorre a produção
de
citocinas
proinflamatórias,
maturação
de
células
dendríticas e outras respostas imunológicas. Os TLRs
também são capazes de reconhecer sinais endógenos
como proteínas do choque térmico e produtos da necrose
celular, assim podem estar envolvidos na auto-imunidade
ou ativação do reparo tecidual.
TLRs são expressos sobre uma variedade de células
do
sistema
imune,
incluindo
macrófagos,
células
dendríticas (DCs), células B, tipos específicos de células T
e até mesmo sobre células não imunológicas como
fibroblastos e células epiteliais. A expressão dos TLRs não
é estática. É modulada rapidamente em resposta a
presença de patógenos, uma variedade de citocinas e o
estresse
ambiental.
Além do mais eles podem ser
expressos intra e extracelularmente. Enquanto certos
16 | P á g i n a
______________Sistema imune inato______________
TLRs (TLRs 1,2,4,5 e 6) são expressos sobre a superfície
celular,
outros
(TLRs
3,7,8
e
9)
são
encontrados
exclusivamente em compartimentos intracelulares como
endossomos,
e
seus
ligantes,
principalmente
ácidos
nucléicos, devem ser internalizados nos endossomos
antes da sinalização ser possível.
Utilizando várias fontes de pesquisa, descreva os principais
TLRs indicando seus locais de expressão e o componente alvo
do seu reconhecimento. Utilize o espaço seguinte para sua
pesquisa.
17 | P á g i n a
______________Sistema imune inato______________
Após a ligação dos TLRs com seus respectivos
componentes alvos, ocorre a ativação de uma cascata
metabólica, que levará a ativação de genes envolvidos na
defesa do hospedeiro conta ação de microrganismos. Para
isso, após a ligação, os TLRs dimerizam-se e submetemse
a
alterações
recrutamento
de
conformacionais
moléculas
requeridas
adaptadoras
para
o
contendo
o
domínio TIR para o domínio TIR dos TLRs. Existem quatro
moléculas
associada
adaptadoras
a
TIR
chamadas
My88D,
proteína
(TIRAP)/My88D-adaptor-like
(MAL),
proteína adaptadora contendo domínio TIR indutora de
IFN-β(TRIF)/molécula 1 contendo domínio TIR (TICAM1),
e molécula adaptadora relacionada a TIRF (TRAM). As
diferentes respostas mediadas por distintos ligantes aos
TLRs podem ser explicadas em parte pelo uso seletivo
dessas
moléculas
adaptadoras.
My88D
e
TIRF
são
responsáveis pela ativação de distintas vias sinalizatórias,
levando à produção de citocinas pró-inflamatórias e IFNs
do tipo I, respectivamente (Figura 2).
Figura 2. Vias de sinalização dos TLRs (modificado de AKIRA et
al., 2006)
18 | P á g i n a
______________Sistema imune inato______________
3.2.3.2 Receptores Scavenger
Os macrófagos possuem receptores glicoprotéicos
transmembranas triméricos identificados como SR-AI, SRAII e MARCO. Esses são denominados como receptores
Scavenger. Eles se ligam a lipídios de baixa densidade,
LPS e ácido lipoteicólico. Sendo assim, estão envolvidos
na detecção de bactérias atuando como receptores para a
fagocitose.
3.2.3.3 Receptores de peptídeos formil
São encontrados na membrana de neutrófilos e
monócitos. Eles ligam-se a formil-meiotionina-leucinafelnilalanina (fMLF) e são em parte responsáveis pela
quimiotaxia de neutrófilos para a área infectada. Eles
também
podem
estar
envolvidos
na
modulação
da
resposta inflamatória através da detecção de ligantes
endógenos para estes recpetores.
3.2.4 Receptores intracelulares
Dois receptores intracelulares têm sido estudados
com grande interesse. O PKR (proteína quinase ativada –
RNA dupla-fita - dsRNA) que reconhece RNA dupla-fita
presente na célula infectada e favorece a ativação do NFκβ (fator nuclear κβ) e do MAP quinase
ína (prote
ativadora da mitose) os quais estão envolvidos na inibição
ou ativação da apoptose em células infectadas por vírus,
ou por células em estresse metabólico.
19 | P á g i n a
______________Sistema imune inato______________
Recentemente,
descobertas
têm
indicado
que
famílias de proteínas com o domínio de oligomerização
ligante de nucleotídeos (NOD) possuem um importante
papel na detecção de bactérias intracelulares. O NOD1 foi
originalmente
identificado
como
uma
molécula
estruturalmente relacionada ao regulador de apoptose
Apaf-1.
Ele
reconhece
diaminopimélico.
Já
o
o
γ-D-glutamil-meso-ácido
NOD2,
mostra
similaridade
estrutural com NOD1, contudo ele reconhece o muramil
dipeptídeo
estruturas
MurNac-L-Ala-pisoGln
reconhecidas
pelo
(MDP).
receptores
NOD
Ambas
estão
presentes nas camadas de PGN (peptídeosglicanos) de
bactérias.
3.3 Citocinas e quimiocinas
Citocinas e quimiocinas são ambas moduladoras das
respostas imune inata e adaptativa. Elas podem ser
produzidas por uma variedade de células incluindo células
do sistema imunológico e células das paredes epiteliais e
endoteliais. Na imunidade inata as principais fontes de
citocinas são os macrófagos, neutrófilos, células natural
killers e keratinócitos. Quimiocinas e citocinas podem
desempenhar
suas
funções
tanto
localmente
como
sistemicamente. O perfil gerado destas substâncias na
resposta imune inata que determinará o balanço Th1 ou
Th2 da resposta imune adquirida. O que demonstra a
ligação entre resposta imune inata e a adquirida.
Pela definição da palavra, citocinas são proteínas
secretadas pelas células do sistema imune inato e
adaptativo, que modulam muitas das funções dessas
células. Já quimiocinas são uma grande família proteínas
20 | P á g i n a
______________Sistema imune inato______________
homólogas estruturalmente das citocinas que estimulam
os movimentos dos leucócitos e regulam a migração dos
leucócitos do sangue para os tecidos.
Citocinas
maturação
das
e
quimiocinas
células
são
dendríticas,
importantes
na
recrutamento
de
células efetoras nos locais de infecção, estimulação da
resposta inflamatória aguda e limitação do espalhamento
do patógeno. Dentre as várias citocinas e quimiocinas
geradas na resposta imune podem-se destacar: TNF-α,
IL-1, IL-4, IL-6, IL-8, IL-12 e os IFNs.
Como forma de escape do sistema imunológico
patógenos
podem
quimiocinas
da
quebrar
resposta
a
rede
de
imunológica
em
citocinas
três
e
vias:
inibição da síntese ou liberação, interferência com os
receptores
de
ligação
ou
pelo
uso
de
subtâncias
antagônicas. Muitos vírus têm a habilidade de inibir as
citocinas pro-inflamatórias ou estimular aquelas antiinflamatórias. É interessante também citar que algumas
bactérias
podem
modular
a
rede
de
citocinas
e
quimiocinas pela produção de proteases que clivam essas
substâncias tornando-as inativas.
O livro Imunologia Celular e Molecular possui um
capítulo que trata apenas das citocinas e quimiocinas. Como
sugestão leia este capítulo.
21 | P á g i n a
______________Sistema imune inato______________
3.4 Células do sistema inume inato
3.4.1 Mucosa epitelial – primeira linha de
defesa
A primeira linha de defesa do organismo contra
invasões de patógenos é representada por barreiras
físicas que impedem os microrganismos de infectar o
ambiente estéril interno do hospedeiro. Fazem parte
desta primeira barreira as superfícies mucosas do trato
respiratório, gastrointestinal, genitário e a pele. Todas
estas barreiras contêm também inúmeras outras formas
de defesa do organismo como células especializadas e
compostos químicos específicos, os quais diferem para
cada localização anatômica.
Os
órgãos
mais
susceptíveis
aos
ataques
de
patógenos são os pulmões e o intestino. Isso se deve ao
fato desses órgãos possuírem uma grande superfície de
contado com o ambiente externo.
Processos fisiológicos como o movimento ciliar,
secreção de uma camada de muco constituído de mucinas
(glicoproteínas), auxiliam na eliminação dos patógenos
que
tentam
se
aderir
às
superfícies
epiteliais
do
hospedeiro. Outras defesas como a secreção de IgA e
lactoferrina também podem inibir essa adesão. Nos
intestinos a flora microbiana comensal auxilia na proteção
inibindo
muitas
microrganismos
vezes
patogênicos
o
desenvolvimento
neste
órgão.
de
Ambiente
desfavorável a proliferação dos patógenos também é
encontrado no estômago, devido a secreção de ácido
clorídrico o qual diminui o pH, provocando a morte dos
agentes infecciosos presentes na lúmen deste órgão.
22 | P á g i n a
______________Sistema imune inato______________
Várias enzimas e peptídeos secretados nas superfícies
mucosas também podem auxiliar a matar microrganismos
invasores. Lisozima é um exemplo de enzima capaz de
degradar a parede celular de bactéria. A lactoferrina
possui grande afinidade por ferro, isso possibilita a
inibição do desenvolvimento de microrganismos que
dependem deste componente para sobrevivência. Já a
lactoperoxidase produz superóxidos que matam bactérias.
Apesar de todos estes mecanismos já citados,
patógenos podem evadir-se e alcançar o ambiente interno
das superfícies epiteliais. Ocorrendo isso, as células
epiteliais possuem mecanismos de percepção da presença
desses microrganismos tanto em suas faces apical, basolateral e intracelular. Vias metabólicas específicas são
ativadas quando receptores ligam-se aos seus ligantes
sobre os microrganismos ou seus produtos. Assim sendo,
ocorre a secreção de citocinas e quimiocinas que atuaram
no recrutamento de tipos celulares que executam a
resposta
imunológica,
assim
como
ativam
fatores
transcripcionais que levarão a vias de resolução da
infecção. Muitas quimiocinas têm sido relacionadas ser
secretadas pelas células epiteliais: IL-8, ENA 78, gro-α,
MIP-2e MCP-1. A interleucina 8 (IL-8) é relacionada com o
recrutamento de neutrófilos para a região baso-lateral.
Monócitos/macrófagos têm seu recrutamento facilitado
pela expressão de receptores ICAM-1 sobre as superfícies
das células.
Peptídeos
como
as
defensinas,
catelicidinas
e
histatinas têm demonstrado ser secretados pelas células
epiteliais e possuem importante papel na defesa do
hospedeiro
(humanos).
As
defensinas
ligam-se
as
membranas de organismos carregados negativamente
23 | P á g i n a
______________Sistema imune inato______________
levando a desestabilização desta. Catelicidinas como as
defensinas podem-se ligar aos LPS e inativam sua função
biolológica. Histatinas estão presentes na saliva humana e
sua ação antimicrobiana é particulamente efetiva contra
fungos.
No intestino, a presença de células epiteliais, células
M e células dendríticas as quais são especializadas na
tomada dos patógenos do lúmen transpondo-os para o
interior do epitélio, em regiões denominadas placas de
Peyer,
facilitam
o
desenvolvimento
da
resposta
imunológica contra estes patógenos. Esta resposta pode
ser dada localmente ou ser amplificada e tornar-se uma
resposta sistêmica.
3.4.2
Células
fagocitárias:
neutrófilos
e
macrófagos
A principal função das células fagocitárias é realizar
a fagocitose dos agentes infecciosos antes que eles
possam espalhar pelo organismo hospedeiro. Uma função
função secundária, porém não menos importante, é
executada
pelos
macrófagos.
Eles
são
capazes
de
funcionar como células apresentadoras de antígenos,
sendo assim, são umas das ligações entre a resposta
imune inata e a resposta imune adaptativa.
As primeiras células reativas ao patógeno invasor
são os macrófagos residentes nos tecidos epiteliais.
Neutrófilos são recrutados posteriormente aos sítios de
infecção pela expressão de citocinas, quimiocinas e sinais
de perigo, que fazem com que estas células abandonem a
corrente sanguínea e migrem para os locais de ação.
24 | P á g i n a
______________Sistema imune inato______________
Tantos macrófagos como neutrófilos possuem muitos
receptores
responsáveis
pelo
reconhecimento
dos
patógenoss. Após fagocitados os microrganismos são
internalizados em fagossomos. Através da atuação de
enzimas contidas nos lisossomos que se fundem ao
fagossomo,
o
conteúdo
fagocitado
é
digerido
por
mecanismos que envolvem reações oxidativas e nãooxidativas.
Os neutrófilos são leucócitos também conhecidos
como polimorfonucleares devido a aparência de seu
núcleo através da microscopia. Seu núcleo é segmentado
em três a cinco porções. Eles possuem grânulos que
contêm agentes microbianos oxidativos e não-oxidativos
que são liberados no fagolisossomo. Entre estes agentes
incluem-se:
defensinas,
proteínas
aumentadora
da
permeabilidade bactericida (B/PI), proteínas catiônicas
que se ligam ao LPS, fosfolipases, lisozimas, lactoferrinas
e catepsina G. Algumas dessas subtâncias também são
importantes
na
regulação
da
resposta
imune.
O
hipoclorito é o principal oxidante bactericida produzido
pelos macrófagos. Ele também é responsável pelo dano
tecidual do hospedeiro. Assim como espécies de oxigênio
reativo
que
são
liberadas
pelo
neutrófilo
e
estão
relacionadas com a inflamação crônica nas quais ocorre a
persistência dos neutrófilos no tecido que foi infectado.
Os macrófagos são originários da hematopoese na
medula óssea. Eles são primeiramente definidos como
monócitos quando ganham a circulação sanguínea ainda
não tendo completado seu desenvolvimento. Uma vez
completado seu desenvolvimento recebem o nome de
macrófago apesar de existirem variações dependendo do
local onde são encontrados. Por exemplo, quando são
25 | P á g i n a
______________Sistema imune inato______________
encontrados no sistema nervoso central recebem o nome
de células da micróglia, quando encontrados nos pulmões,
macrófagos alveolares; no fígado, células de Kupffer; e na
medula
quando
apresentam
múltiplos
núcleos,
osteoclastos. Como os neutrófilos os macrófagos podem
responder a presença de patógenos nos sítios de infecção.
Contudo eles apresentam a característica de não serem
completamente
diferenciados,
o
que
possibilita
sua
divisão nos sítios de infecção possibilitando sua maior
permanência durante a inflamação. Os macrófagos são
principais células nos estágios precoces da infecção.
3.4.3 Células dendríticas (DCs)
Entre os componentes celulares identificados no
sistema imune inato as células dendríticas têm sido
identificadas
como
o
principal
tipo
celular
para
apresentação de antígenos, sendo então classificadas
como células apresentadoras profissionais. Estas células
são capazes de reconhecer muitos padrões antigênicos
através de vários receptores padrões de reconhecimento
expressos por elas como os TLRs. Tem sido demonstrado
que células dendríticas estão presentes em quase todos
órgãos periféricos e tecidos linfóides, fato este que
contribui para sua ação contra agentes infecciosos, pois
virtualmente elas estão presentes em todos possíveis
sítios de entrada dos antígenos. Recentemente têm-se
sugerido que estas células também estão envolvidas em
processos
de
indução
e
manutenção
da
tolerância
antigênica.
26 | P á g i n a
______________Sistema imune inato______________
O ciclo de vida das células dendríticas inicia com as
células
precursoras
(incluindo
monócitos),
as
quais
circulam na circulação sanguínea e são continuamente
geradas pelos progenitores na medula óssea. A migração
para órgãos não linfóides induz a diferenciação das células
precursoras das células dendríticas em células dendríticas
que tornam-se residentes nos tecidos celulares (interstício
dos órgãos periféricos e pele). Contudo, estas células
encontram-se nesses locais em um estado (fenótipo)
imaturo.
Sua
maturação
ocorre
através
do
reconhecimento de PAMPs por estas células somados a
um ambiente ideal de moléculas (citocinas, quimiocinas,
DAMPs, etc) que auxiliam no seu desenvolvimento. A
ativação das células dendríticas leva a apresentação de
antigênica através de moléculas do complexo principal de
histocompatibilidade da classe II (MHC-II) de peptídeos
derivados do processamento do antígeno fagocitado.
Células dendríticas maturas são capazes de secretar
uma variedade de citocinas, dentre elas IL-12p40 e p70,
TNF-α, IL-1β, IL-6 e óxido nítrico. Estas citocinas estão
envolvidas
no
desenvolvimento
e
manutenção
de
processos inflamatórios relacionados à resposta resposta
imune inata. Ela também secreta a IL-2 que é uma
molécula chave na indução da imunidade T dependente
(imunidade adquirida) ou na efetivação de mecanismos
tolerogênicos.
O processo de maturação das células dendríticas,
após ativado, faz com que estas células alterem o padrão
de moléculas receptoras de quimiocinas. Sendo assim,
elas adquirem a capacidade de migrar dos tecidos
periféricos para os vasos linfáticos onde ganham a
circulação linfática e posteriormente os linfonodos, onde
27 | P á g i n a
______________Sistema imune inato______________
atuaram como apresentadoras de antígeno estimulando a
resposta imune adquirida. Para isso ocorre uma regulação
negativa para a expressão de CCR1 e CCR5, e positiva
para CCR7 o qual direciona as DCs via quimiocinas CCL19
e CCL21.
Figura 3. Revisão das DCs e suas principais
características como células apresentadoras de antígenos
(modificado de ADAMS et al., 2005).
3.4.4 Células natural killers (NK)
Células natural killers são derivadas dos mesmos
precursores linfóides de linfócitos T. Essas células contêm
importantes moléculas chamadas perforinas e granzimas,
as quais são liberadas durante o contato célula-célula e
são responsáveis por matar células tumorais e aquelas
infectadas por vírus. Células NK também podem atuar
através da via Fas/FasL e são a principal fonte de IFN-γ
que ativa os macrófagos para matar patógenos. Já os
macrófagos secretam IL-12 que é essencial para a
atividade citotóxica das células natural killers.
28 | P á g i n a
______________Sistema imune inato______________
Diferentemente de células T citotóxicas, células NK
não reconhecem especificamente os antígenos, seus
receptores não são codificados por genes que sofrem
recombinação somática que determinam a especificidade,
assim elas diferem das células relacionadas ao sistema
imune adaptativo, representando um braço da resposta
imune inata. Contudo, quando analisadas as bases de
suas relações de linhagem, repertório de receptores e
funções efetoras, células NK parecem ser um tipo células
que interliga os sistemas imune inato e adaptativo.
Através do reconhecimento dos patógenos por seus
receptores elas respondem rapidamente ativando sua
funções efetoras. Uma variedade de receptores são
expressos pelas células naturais killers, dentre estes
destacam-se CD2, CD69, CD226 (DNAM-1), CD44 e Ly6,
alguns destes envolvidos com a atividade citotóxica da
células NK e outros envolvidos com a produção de
citocinas que
atuam como reguladoras
da resposta
imunológica. Uma família de receptores denominada KIR
(receptores
funções
parecidos
ativadoras
e
com
imunoglobulinas)
inibitórias.
O
KIR
possui
inibitório
reconhece células que expressam níveis normais de MHCI protegendo-as da função lítica das NKs. Células que
expressam baixa quantidade de MHC-I são então lisadas.
Já o receptor KIR ativador possui a função de regular as
funções das células NKs.
Alguns estudos citam sobre a possibilidade que
células NK podem interagir diretamente com células T
CD4+. Isso é sugerido pelo fato das células NK humanas
possuirem MHC-II. Estudos recentes têm demonstrado
que a ativação de células NK humanas por citocinas
“inatas” (IL-12, IL-15) é suficiente para a expressão de
29 | P á g i n a
______________Sistema imune inato______________
CD86 sobre essas células. Assim sendo, a expressão do
CD86 que é o ligante do CD28 (potente receptor
coestimulatório) possibilita a ativação de células T CD4+,
o que demonstra a interligação entre os sistemas inato e
adaptativo através das células NK.
Existem um pequeno número de células do sistema
imune que compartinham características com as células
naturais killers e as células T. Essas são denominadas
células
NK-T.
sublinhagem
Elas
constituem
compropriedades
uma
conservada
únicas,
incluindo
reatividade para glicolipídeos apresentado por Cd1d,
expressão de um receptor células T (TCR) cadeia α, e um
requerimento não usual para seleção tímica. Essas células
produzem
rapidamente
citocinas
que
influenciam
a
resposta imunológica e processos patogênicos.
3.4.5 Mastócitos
Células
medula
mastócitos
óssea.
Elas
(MCs)
circulam
têm
no
sua
origem
sangue
em
na
baixa
concentração e são encontradas em todos os tecidos, mas
são
especialmente
abundantes
nos
limites
entre
o
organismo e o ambiente, como nas mucosas das vias
aéreas e intestino, e na pele. Este tipo celular possui uma
espectativa de vida de 1 a 6 meses.
MCs respondem diretamente e indiretamente aos
patógenos, tornando estas células importantes sentinelas
do
sistema
imunológico.
Elas
podem
então
iniciar
mecanismos importantes da resposta imune inata ao
patógeno. Entretanto, células mastócitos também podem
trabalhar como influenciadoras de uma boa resposta
30 | P á g i n a
______________Sistema imune inato______________
adaptativa, pois contribuem com a maturação, migração e
função das células dendríticas, além de interagirem com
os linfócitos T e B.
Como
células
do
sistema
inume
inato,
MCs
produzem importantes compostos com função bactericida
como
as
catelicidinas,
inflamatórias
como
além
heparina,
de
substâncias
histamina
e
pro-
fatores
quimiostáticos para eosinófilos. Elas também podem
contribuir no controle da reposta através da ação de
proteases que degradam mediadores endógenos, os quais
são super-regulados durante a infecção. A resposta das
células matócitos não é restringida às bactérias, elas
também são reativas a muitos patógenos como parasitas
e vírus.
3.5 Conclusão
O sistema inume inato constitui a primeira linha
defesa
do
organismo,
através
de
barreiras
físicas,
químicas e elementos celulares. A interação entre estes
componentes possibilita o reconhecimento e muitas vezes
a eliminação do patógeno invasor através de mecanismos
próprios. Como resultado do reconhecimento, o sistema
imune inato é capaz de influenciar outro mecanismo
imunológico do hospedeiro, conhecido como resposta
imune
adaptativa.
Existem
vários
componentes
do
sistema imune inato que são importantes para esta
interação, contudo três componentes se destacam, sendo
eles, os TLRs, as células dendríticas e as citocinas. Esta
interação entre dois braços da reposta imunológica
possibilita
o
organismo
do
hospedeiro
combater
eficientemente o patógeno.
31 | P á g i n a
______________Sistema imune inato______________
3.6 Bibliografia
ABBAS, A. K.; LICHTMANA, A. H.; PILLAI, S. Imunologia
Celular e Molecular. Rio de Janeiro: Elsevier, 6ed. 2008.
ADAMS, S.; O’NEILL, D. W.; BHARDWAJ, N. Recent
Advances in Dendritic Cell Biology. Journal of Clinical
Immunology, v.25, n.2, p.87-98, 2005.
AKIRA,
S.;
UEMATSU,
S.;
TAKEUCHI,
O.
Pathogen
Recognition and Innate Immunity. Cell, v.124, p.783–
801, 2006.
ALBERTS,
B.;
JOHNSON,
A.;
WALTER,
P.
Biologia
molecular da célula. São Paulo: Artmed, 4ed. 2004.
BASSET, C.; HOLTON, J.; O’MAHONY, R.; ROITT, I. Innate
immunity and pathogen–host interaction. Vaccine, v.21,
p.s2/12-s2/23, 2003.
FILHO, G. B. Bogliolo Patologia Geral. Belo Horizonte:
Guanabara Koogan, 3ed. 2003.
JANEWAY, JR. C. A.; MEDZHITOV, R. Innate immune
recognition. Annu. Rev. Immunol., v.20, p. 197-216,
2002.
LEE, H. K.; IWASAKI, A. Innate control of adaptive
immunity:
Dendritic
cells
and
beyond.
Seminars
in
Immunology, v.19, p.48-55, 2007.
MEDZHITOV, R. Toll-like receptors and innate immunity.
Nature reviews Immunology, v.1, p.135-145, 2001.
MEDZHITOV, R.; JANEWAY JR., C. A. Innate immunity:
impact
on
the
adaptive
immune
response.
Current
Opinion in Immunology, v.9, p.4-9, 1997.
32 | P á g i n a
______________Sistema imune inato______________
TEGNÉR, J.; NILSSON, R.; BAJIC, V. B.; BJÖRKEGREN, J.;
RAVASI, T. Systems biology of innate immunity. Cellular
Immunology, v.244, p.105–109, 2006.
TURVEY, S. E.; BROIDE, D. H. Innate immunity. American
Academy of Allergy, Asthma and Immunology. p.s24-s32,
2010.
WILSON, N. S.; VILLADANGOS, J. A. Lymphoid organ
dendritic cells: beyond the Langerhans cells paradigm.
Immunology and Cell Biology, v.82, p.91-98, 2004.
WONG, S. Y. C. Innate immune trouble detectors. Trends
in Immunology, v.22, n.5, p.235-236, 2001.
33 | P á g i n a