O Músculo
João Ricardo Paulo Ferreira, 4/Novembro/2012
Cruz dos Morouços
Índice
Funções do sistema muscular……………………………………………………………………………………..2
Características gerais do funcionamento do músculo…………………………………………………..2
Propriedades do músculo…………………………………………………………………………………………….2
Tipos de tecido muscular……………………………………………………………………………………………..3
Estrutura do músculo esquelético……………………………………………………………………………….4
Tecido conjuntivo…………………………………………………………………………………………………………5
Nervos e vasos sanguíneos…………………………………………………………………………………………..5
Fibras musculares…………………………………………………………………………………………………………6
Modelo do deslizamento dos filamentos……………………………………………………………………..9
Fisiologia das fibras do músculo esquelético…………………………………………………………….10
Potenciais de membrana……………………………………………………………………………………………10
Canais iónicos…………………………………………………………………………………………………………….11
Potenciais de ação……………………………………………………………………………………………………..11
Junção neuromuscular……………………………………………………………………………………………….13
Acoplamento, excitação, contração……………………………………………………………………………15
Movimento das pontes………………………………………………………………………………………………17
Relaxamento muscular……………………………………………………………………………………………….18
Fisiologia do músculo esquelético…………………………………………………………………………….19
Contração muscular……………………………………………………………………………………………………19
Intensidade do estímulo e contração muscular………………………………………………………….20
Frequência do estímulo e contração muscular……………………………………………………………22
Tipos de contração muscular…………………………………………………………………………………….23
Comprimento vs. Tensão……………………………………………………………………………………………24
Fadiga………………………………………………………………………………………………………………………..25
Fontes de energia………………………………………………………………………………………………………26
Creatina fosfato………………………………………………………………………………………………………….26
Glicólise……………………………………………………………………………………………………………………..26
Respiração anaeróbia…………………………………………………………………………………………………31
Respiração aeróbia…………………………………………………………………………………………………….32
Fibras lentas e fibras rápidas…………………………………………………………………………………….36
1
Funções do Sistema Muscular
A maior parte dos movimentos no nosso corpo são realizados através de contrações
musculares, desde o bater do coração ao correr a maratona.
As principais funções dos músculos são:
Movimento corporal;
Manutenção da postura;
Respiração;
Produção de calor corporal;
Comunicação;
Contrição de órgãos e vasos;
Batimento cardíaco.
Características Gerais do Funcionamento do Músculo
Propriedades do Músculo
O músculo tem quatro características funcionais fundamentais:
Contratilidade (capacidade que um músculo tem de para se contrair,
produzindo uma determinada força);
Excitabilidade (capacidade que um músculo tem para responder a um
estímulo);
Extensibilidade (o músculo pode ser estirado até ao seu comprimento normal
em repouso e, em dado grau, para lá desse comprimento);
Elasticidade (depois de serem estirados, os músculos retornam ao seu
comprimento em repouso original).
2
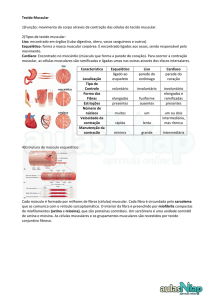
Tipos de Tecido Muscular
Músculo
Esquelético
Músculo Liso
Músculo
Cardíaco
Localização
Inserido nos ossos
- Paredes dos
órgãos ocos
- Vasos sanguíneos
- Olhos
- Glândulas
- Pele
Coração
Forma das células
Muito longas e
cilíndricas
Em forma de fusos
Cilíndricas e
ramificadas
Núcleo
Múltiplos, com
localização
periférica
Único com
localização central
Único com
localização central
As fendas
sinápticas juntam
diversas células do
músculo liso
visceral.
Os discos
intercalares unem
as células umas às
outras
Características
Características
especiais das
junções celulares
Estrias
Sim
Não
Sim
Controlo
Voluntário e
involuntário
(reflexos)
Involuntário
Involuntário
Capacidade de
controlo
espontâneo
Não
Sim
Sim
Movimento
corporal
Mobilização dos
alimentos no tubo
digestivo,
esvaziamento da
bexiga, regulação
do diâmetro dos
vazos sanguíneos,
alteração do
tamanho da pupila,
contração de
muitos canais
glandulares,
movimento dos
pelos e muitas
outras funções
Bombeia o sangue:
as contrações
constituem a
principal força para
impulsionar o
sangue nos vasos
sanguíneos
Função
3
Estrutura do Músculo Esquelético
Os músculos esqueléticos são constituídos por fibras musculares esqueléticas,
associadas a pequenas quantidades de tecido conjuntivo, vasos sanguíneos e nervos.
Cada fibra muscular esquelética é uma célula cilíndrica única com diversos núcleos
localizados à sua periferia, junto da membrana celular.
Estas fibras desenvolvem-se a partir de células multinucleadas pouco diferenciadas
denominadas mioblastos. Os mioblastos convertem-se em fibras musculares quando
as proteínas contrácteis se acumulam no seu citoplasma.
O número de fibras musculares esqueléticas mantém-se relativamente constante após
o nascimento. Por isso, a hipertrofia muscular após o nascimento resulta de um
aumento do tamanho das fibras muscular do indivíduo. Desta forma, a hipertrofia
muscular em resposta ao exercício é mais devida a um aumento do tamanho das fibras
musculares do que ao aumento do seu número.
4
Tecido Conjuntivo
Envolvendo cada fibra muscular existe uma delicada lâmina externa composta,
essencialmente, por fibras reticulares. Esta lâmina externa é produzida pela fibra
muscular, o sarcolema. O endomísio, rede delicada de tecido conjuntivo laxo, com
numerosas fibras reticulares, envolve cada fibra muscular por fora da lâmina externa.
As fibras musculares com o seu endomísio são envolvidas por outra camada de tecido
conjuntivo mais denso, o perimísio. Cada conjunto de fibras envolvido pelo perimísio é
um feixe muscular. Um músculo é constituído por muitos feixes agrupados e rodeados
por uma terceira camada, mais espessa, o epimísio, formado por tecido conjuntivo
denso, fibroso e colagénico e que cobre toda a superfície muscular.
Uma fáscia consiste em tecido conjuntivo fibroso que envolve o corpo, formando uma
bainha sob a pele. Também separa cada músculo e, em alguns casos, envolve grupos
musculares. A fáscia em torno de um músculo chama-se epimísio.
O tecido conjuntivo do músculo mantém juntas as células musculares e liga os
músculos aos tendões ou insere-os nos ossos.
Nervos e Vasos Sanguíneos
São muitos os nervos e vasos sanguíneos que se estendem para os músculos
esqueléticos.
Neurónios motores – são células nervosas especializadas em que os seus
corpos celulares se localizam no tronco cerebral ou na medula espinhal e os
axónios estendem-se atá às fibras musculares esqueléticas através dos nervos.
Os neurónios motores estimulam a contração muscular e enervam mais do que
uma fibra muscular.
Juntamente com o nervo segue também uma artéria e uma ou duas veias, até
às camadas de tecido conjuntivo do músculo esquelético.
5
Fibras Musculares
Os múltiplos núcleos de cada fibra muscular encontram-se imediatamente sob o
sarcolema, enquanto a maior parte do interior da fibra é constituída por miofibrilhas.
Entra as miofibrilhas estão alojados outros organelos, como numerosas mitocôndrias e
grânulos de glicogénio. O citoplasma sem as miofibrilas chama-se sarcoplasma.
Miofibrilhas:
Estrutura filamentosa que se estende de uma extremidade do músculo para a
outra;
Compõem-se por duas espécies de filamentos proteicos chamados
miofilamentos:
Miofilamentos de actina;
Miofilamentos de miosina.
Os miofilamentos de actina e miosina organizam-se em unidades altamente
organizadas chamadas sarcómeros, que se unem topo a topo para formar miofibrilhas.
6
Miofilamentos de actina e miosina
Actina
Cada miofilamento de actina é composto por:
Duas cadeias de actina fibrosa (actina F);
Uma série de moléculas de tropomiosina;
Uma série de moléculas de troponina.
As duas cadeias de actina F enrolam-se numa dupla hélice que se estende a todo o
comprimento do miofilamento de actina. Cada cadeia de actina F é um polímero de
pequenas unidades globulares chamadas monómeros de actina globular (actina G).
Cada monómero de actina G tem um local específico ondem se podem ligar moléculas
de miosina durante a contração muscular.
A tropomiosina é uma proteína alongada inserida na fenda da dupla hélice da actina F.
Cada molécula de tropomiosina é suficientemente longa para cobrir sete locais ativos
de actina G.
A troponina é constituída por três subunidades:
Uma que se liga à actina;
Uma que se liga à tropomiosina;
Uma que se liga a iões cálcio.
As moléculas de troponina encontram-se entre as extremidades das moléculas de
tropomiosina, nos sulcos entre as cadeias de actina F.
O complexo de tropomiosina e troponina regula a interação entre os locais ativos da
actina G e a miosina.
Miosina
Os miofilamentos de miosina compõem-se de duas moléculas de miosina alongadas,
com a forma de um taco de golfe. Cada molécula de miosina consiste em:
Duas moléculas de miosina pesada, que se juntam de modo a formar uma
porção cilíndrica paralela ao miofilamento de actina;
Duas cabeças que se estendem lateralmente.
Quatro cadeias de miosina leve ligam-se às cabeças de cada molécula de miosina.
As cabeças de miosina têm três propriedades importantes:
Podem ligar-se a sítios ativos nas moléculas de actina, formando pontes
cruzadas ou, simplesmente, pontes.
A cabeça liga-se à porção cilíndrica da miosina por uma zona encurvada que se
pode dobrar e estreitar durante a contração. Uma segunda zona curva fica a
curta distância da cabeça na porção cilíndrica da molécula de miosina.
As cabeças têm atividade de ATPase, atividade enzimática que desdobra a
adenosina trifosfato (ATP), libertando energia. Parte desta energia é usada para
dobrar a região em charneira da molécula de miosina durante a contração.
7
Sarcómeros
Cada sarcómero estende-se de uma linha Z para a linha Z imediatamente a seguir. A
linha Z é uma rede filamentosa de proteínas que forma uma estrutura em forma de
disco, que faz a ligação dos miofilamentos de actina. O arranjo dos miofilamentos de
actina e dos miofilamentos de miosina dá à miofibrilha uma aparência estriada quando
vista longitudinalmente.
Cada banda I, ou isotrópica (banda clara), inclui uma linha Z e estende-se de cada lado
dessa linha para as extremidades dos miofilamentos de miosina.
Cada banda A, ou anisotrópica (banda escura), estende-se ao comprimento dos
miofilamentos de miosina num sarcómero. Os miofilamentos de miosina e actina
sobrepõem-se em parte da sua extensão em ambas as extremidades da banda A.
No centro de cada banda A está uma pequena banda chamada zona H, onde os
miofilamentos de actina e miosina não se sobrepõem e apenas estão presentes
miofilamentos de miosina.
No meio da zona H encontra-se uma linha escura chamada linha M que consiste em
delicados filamentos que se ligam ao centro dos miofilamentos de miosina e os
mantém no local.
8
Modelo do Deslizamento dos Filamentos
1. Os miofilamentos de actina e miosina num músculo relaxado e num músculo
contraído têm o mesmo comprimento. Os miofilamentos não mudam de
comprimento durante a contração do músculo esquelético.
2. Durante a contração, os miofilamentos de actina em cada extremidade do
sarcómero deslizam sobre os miofilamentos de miosina, na direção uns dos
outros. Em consequência, os discos Z aproximam-se e o sarcómero encurta-se.
3. Quando os miofilamentos de actina deslizam sobre os miofilamentos de
miosina, as zonas H e as bandas I tornam-se mais estreitas. As bandas A, que
têm um comprimento igual ao dos miofilamentos de miosina, não se estreitam,
porque o comprimento dos miofilamentos de miosina se mantém.
4. Num músculo totalmente contraído, as extremidades dos filamentos de actina
sobrepõem-se e a zona H desaparece.
9
Fisiologia das Fibras do Músculo Esquelético
Os axónios das células nervosas estendem-se do encéfalo e da medula espinhal para as
fibras dos músculos esqueléticos. O sistema nervoso controla as contrações dos
músculos esqueléticos através de sinais elétricos, chamados potenciais de ação, que
são transmitidos ao longo dos neurónios até às fibras musculares. Os potenciais de
ação transmitidos pelos neurónios estimulam a produção de potenciais de ação nas
fibras musculares, que as levam a contrair.
Potenciais de Membrana
As membranas celulares são polarizadas, o que significa que existe uma diferença de
carga elétrica através da membrana, antes de poderem ser gerados potenciais de
ação. Esta diferença de carga chama-se potencial de repouso. A carga negativa na face
interior da membrana celular, em comparação com a da face exterior, resulta
principalmente das diferenças de concentração de iões e moléculas carregadas
através da membrana celular e das suas características de permeabilidade. A
concentração de iões K+ no interior da célula é muito superior à sua concentração no
exterior da célula. A membrana celular é relativamente permeável aos iões K+ e muito
menos permeável a moléculas com carga negativa que se encontram no interior da
célula. Consequentemente, os iões K+ com carga positiva tendem a difundir-se para
fora da célula, deixando para trás as moléculas com carga negativa. A membrana fica
polarizada quando a tendência para os iões K+ se difundirem para fora da célula é
resistida pelas cargas negativas das moléculas dentro da célula.
As diferenças de potencial através das membranas das células nervosas e das fibras
musculares estão entre os -70 e -90mV. A diferença de potencial traduz-se num
número negativo porque a superfície interior da membrana é negativa em
comparação como exterior.
10
Canais Iónicos
A difusão de iões, aquando de uma estimulação, através de canais iónicos, altera as
suas cargas através da membrana celular e produz um potencial de ação.
O potencial de ação é o reverso do potencial de repouso, no sentido em que o interior
da membrana é positivamente carregado em relação ao exterior.
Há dois tipos de canais iónicos responsáveis por produzir potenciais de ação:
Canais iónicos com portão de ligando – Canais que abrem em resposta à
ligação do ligando a um recetor que é parte do canal iónico.
Canais iónicos com portão de voltagem – Abrem e fecham em resposta a
pequenas alterações (carga) através da membrana celular.
Os canais iónicos com portão de ligando ou com portão de voltagem são específicos
para o tipo de ião que neles passa. O tipo específico de canais iónicos que se abrem
determina que iões se deslocam através da membrana.
Potenciais de Ação
O potencial de ação leva de 1 a poucos milésimos de segundo a ocorrer e tem duas
fases, designadas por despolarização e repolarização.
Despolarização – Dá-se quando o interior da membrana celular se torna
menos negativo, o que é indicado pelo movimento ascendente da curva até
atingir o zero. A fase de despolarização do potencial de ação é desencadeada,
se a despolarização alterar o potencial da membrana para um valor chamado
limiar de estimulação (-55mV).
Repolarização – Regresso do potencial de membrana ao valor de repouso.
11
Como acontece?
A despolarização e a repolarização resultam da abertura e encerramento dos canais
iónicos com portão. Antes de uma célula nervosa ou muscular ser estimulada, esses
canais estão fechados. Quando a célula é estimulada, os canais de Na + abrem e o Na +
difunde-se para dentro da célula. Os iões de Na +, com carga positiva, tornam o interior
da membrana celular menos negativo. Se a despolarização atingir o limiar, muitos
canais de Na+ com portão abrem-se rapidamente e ou iões Na+ difundem-se para o
interior da célula, até que este fique positivo por um curto período de tempo. As
alterações subsequentes da permeabilidade da membrana celular interrompem a
despolarização e iniciam a repolarização, pouco depois de o interior da membrana se
tornar positivo. A fase de repolarização resulta do encerramento dos canais de Na +
com portão e da abertura dos canais de K+ com portão. Assim, cessa o movimento do
Na+ para dentro da célula e aumenta o movimento do K+ para fora da célula. Estas
alterações fazem com que o interior da membrana celular se torne mais negativo e o
exterior mais positivo. O potencial de ação termina e o potencial de membrana em
repouso é restabelecido quando se encerram os canais de K+ com portão.
Princípio do Tudo-ou-nada
Os potenciais de ação ocorrem de acordo com o princípio do tudo-ou-nada.
Parte do “tudo” - Se um estímulo é suficientemente forte para produzir uma
despolarização que atinja o limiar ou mesmo que o ultrapasse, todas as
alterações de permeabilidade responsáveis por um potencial de ação se
sucedem sem parar e são de magnitude constante.
Parte do “nada” – Se o estímulo é tão fraco que a despolarização não atinge o
limiar, são poucas as alterações da permeabilidade. O potencial da membrana
regressa ao nível de repouso, após um brave período sem produzir um
potencial de ação.
Frequência do potencial de ação – número de potenciais de ação produzidos por
unidade de tempo. À medida que aumenta a força do estímulo aplicada à célula
nervosa ou muscular, uma vez atingido o limiar, a frequência do potencial de ação
aumenta.
12
Junção Neuromuscular
Os axónios dos neurónios motores transportam potenciais da ação a alta velocidade,
do encéfalo e medula espinal para as fibras musculares esqueléticas. Os axónios
ramificam-se repetidamente e cada ramo projeta-se para uma única fibra muscular
para a inervar. Assim, cada fibra muscular recebe o ramo de um axónio e cada axónio
inerva mais de uma fibra muscular.
Junto de cada fibra muscular, cada ramo axonal forma um terminal nervoso alargado
que se aloja numa invaginação do sarcolema formando uma sinapse, ou junção
neuromuscular, que consiste nas terminações axonais e na área de sarcolema da fibra
muscular que inervam. Cada axónio acaba no terminal pré-sináptico. O espaço entre
este e a fibra muscular é a fenda sináptica e a membrana celular muscular na área de
junção é a placa motora ou membrana pós-sináptica.
13
Cada terminal pré sináptico contém dentro da sua membrana muitas vesículas
esféricas, chamadas vesículas sinápticas. As vesículas contêm acetilcolina (ACh),
molécula orgânica composta por ácido acético e colina, que atua como
neurotransmissor.
Um neurotransmissor é uma substância libertada por uma membrana pré-sináptica
que se difunde através de uma fenda sináptica e estimula (ou inibe) a produção de um
potencial de ação na membrana pós-sináptica.
Como acontece?
Quando o potencial de ação atinge o terminal pré-sináptico provoca a abertura dos
canais de portão de voltagem do cálcio (Ca2+) na membrana celular do axónio, o que
leva à difusão dos iões Ca2+ para dentro da célula. Uma vez dentro da célula, os iões
Ca2+ provocam uma secreção do conteúdo de diversas vesículas sinápticas, por
exocitose, do terminal pré-sináptico para o interior da fenda sináptica. As moléculas de
acetilcolina libertadas das vesículas sinápticas difundem-se, depois, através da fenda e
ligam-se a moléculas recetoras localizadas na membrana pós-sináptica. Isto leva à
abertura de canais de Na+ com portão de ligando, aumentando a permeabilidade da
membrana aos iões de sódio (Na+). O Na+ difunde-se então para dentro da célula,
produzindo despolarização da membrana. No músculo esquelético, cada potencial de
ação do neurónio motor causa uma despolarização que excede o limiar, resultando na
produção de um potencial de ação na fibra muscular.
Na fenda sináptica, a acetilcolina desdobra-se rapidamente em ácido acético e colina,
por ação da acetilcolinesterase. A colina é reabsorvida pelo terminal pré-sináptico e
combina-se com ácido acético produzido na célula para formar mais acetilcolina, que
entre nas vesículas sinápticas. O ácido acético é um intermediário no processo do
metabolismo e pode ser captado e utilizado por diversas células depois de se difundir a
partir da área da junção neuromuscular.
14
Acoplamento, Excitação, Contração
O mecanismo pela qual um potencial de ação leva à contração da fibra muscular
chama-se acoplamento excitação contração e envolve o sarcolema, os túbulos T, o
retículo endoplasmático, o Ca2+ e a troponina.
Túbulos T
O sarcolema tem ao longo da sua superfície muitas invaginações tubulares chamadas
túbulos T ou transversais. Os túbulos T projetam-se para dentro das fibras musculares
e enrolam-se em torno dos sarcómeros, na região onde os miofilamentos de actina e
miosina se sobrepõem.
Retículo sarcoplasmático
Suspenso no sarcoplasma, entre os túbulos T, está um retículo endoplasmático liso
altamente especializado, que se chama retículo sarcoplasmático. Perto dos túbulos T,
o retículo sarcoplasmático alarga-se de modo a formar cisternas terminais. O conjunto
de um túbulo T e duas cisternas terminais adjacentes chama-se tríade. O retículo
sarcoplasmático transporta ativamente iões de cálcio para dentro do lúmen (as
concentrações de iões cálcio no retículo sarcoplasmático são 2000 vezes superiores
que do sarcoplasma do músculo em repouso).
15
Como acontece?
O mecanismo acoplamento excitação contração começa na junção neuromuscular,
com a produção de um potencial de ação no sarcolema. Esse potencial de ação
propaga-se ao longo de todo o sarcolema da fibra muscular. Quando o potencial de
ação atinge os túbulos T, as membranas destes sofrem despolarização, porque os
túbulos T são invaginações do sarcolema. Os túbulos T transportam a despolarização
para o interior da fibra muscular. Uma vez que os túbulos T atingem a área das tríades,
a sua despolarização leva à abertura dos canais de Ca2+ com portão de voltagem.
Quando os canais de Ca2+ do retículo sarcoplasmático se abrem, os iões Ca2+ difundemse rapidamente do retículo para o sarcoplasma que rodeia as miofibrilhas.
Os iões Ca2+ ligam-se à troponina dos miofilamentos de actina. A combinação dos iões
Ca2+ com a troponina faz com que o complexo troponina-tropomiosina se afunde e
rode no espaço entre as duas moléculas de actina F, expondo assim os locais ativos nos
miofilamentos de actina. Estes locais expostos ligam-se às cabeças das moléculas de
miosina para formar pontes. O movimento das pontes leva à contração.
16
Movimento das Pontes
As moléculas das cabeças da miosina movem-se na região da charneira da actina
forçando-a a deslizar à superfície do miofilamento de miosina. Após este movimento,
cada cabeça de miosina liberta-se da actina e retorna à posição original, podendo
então formar outra ponte num local diferente do miofilamento de actina (ciclo de
formação, movimento e libertação).
Como acontece?
A energia de uma molécula de ATP é necessária para cada ciclo de formação,
movimento e libertação da cabeça da molécula de miosina.
Por ação da ATPase da cabeça do miofilamento de miosina, o ATP desdobra-se em ADP
e uma molécula de fosfato, sendo a energia resultante desta reação armazenada na
cabeça. Tanto o ADP como o fosfato permanecem ligados à cabeça de miosina.
Em consequência do desdobramento do ATP, a ponte liberta-se e a cabeça retorna à
sua posição original. Então, a molécula de miosina liga-se a outro sítio ativo da actina
para formar uma nova ponte e o fosfato é libertado da cabeça de miosina.
Muita da energia armazenada é utilizada para formação e movimento das pontes e a
molécula de ADP é então libertada da cabeça de miosina. Antes da ponte poder ser
libertada para um novo ciclo, é necessário que uma nova molécula de ATP se ligue à
cabeça da molécula de miosina.
O movimento das moléculas de miosina na formação de pontes chama-se choque de
potência, enquanto que o retorno da cabeça de miosina à sua posição original após a
libertação da ponte se chama choque de recuperação.
17
Relaxamento Muscular
O relaxamento muscular resulta do transporte ativo de iões Ca 2+ de volta ao retículo
sarcoplasmático. À medida que diminui a concentração de iões Ca 2+ no sarcoplasma,
estes difundem-se a partir das moléculas de troponina. O complexo troponinatropomiosina restabelece, então, a sua posição, o que bloqueia as locais ativos das
moléculas de actina. Em consequência, as pontes não se podem reformar depois de
terem sido libertadas e dá-se o relaxamento muscular.
Além da energia necessária para a contração muscular, é também necessária energia
para o relaxamento. O transporte ativo de iões Ca2+ para o interior do reticulo exige
ATP. Os processos de transporte ativo que mantêm as concentrações normais de Na + e
K+ através do sarcolema exigem também ATP. A quantidade de ATP necessária à
formação de pontes, durante a contração, é muito maior do que as outras
necessidades energéticas do músculo esquelético.
18
Fisiologia do Músculo Esquelético
Contração Muscular
Encurtamento de um músculo em resposta a um estímulo que causa um potencial de
ação em uma ou mais fibras musculares.
Fases da contração muscular:
Fase de latência ou demora;
Fase de encurtamento;
Fase de relaxamento.
Fase de latência
Um potencial de ação propaga-se para o terminal pré-sináptico do neurónio
motor.
O potencial de ação causa o aumento da permeabilidade do terminal présináptico.
Os iões de cálcio difundem-se para o terminal pré-sináptico, fazendo com que a
acetilcolina contida em diversas vesículas sinápticas seja libertada por
exocitose.
A acetilcolina libertada do terminal pré-sináptico difunde-se através da fenda
sináptica e liga-se às moléculas recetoras da acetilcolina na membrana póssináptica do sarcolema.
A combinação da acetilcolina ao seu local recetor faz abrir os canais de Na+ com
portão de ligando e a membrana pós-sináptica torna-se mais permeável a estes
iões.
Os iões de sódio difundem-se para a fibra muscular, causando uma
despolarização local que excede o limiar de estimulação e produz um potencial
de ação.
A acetilcolina é rapidamente degradada, na fenda sináptica, em ácido acético e
colina, limitando assim o período de tempo em que a acetilcolina está ligada ao
recetor. O resultado é que um potencial de ação pré-sináptico produz apenas
um potencial de ação pós-sináptico nas fibras musculares.
O potencial de ação produzido na fibra muscular propaga-se da membrana póssináptica, localizada perto do meio da fibra, para ambas as extremidades e para
dentro dos túbulos T.
A despolarização que se dá nos túbulos T em resposta ao potencial de ação faz
abrir os canais de Ca2+ com portão de voltagem da membrana do retículo
sarcoplasmático, ficando esta muito permeável aos iões de cálcio.
Os iões de cálcio difundem-se do retículo sarcoplasmático para o sarcolema.
19
Os iões de cálcio ligam-se á troponina; o complexo troponina-tropomiosina
altera a sua posição e expõe o local de ligação nos miofilamentos de actina.
Fase de encurtamento
As pontes entre as moléculas de actina e as moléculas de miosina formam-se,
movem-se, libertam-se e voltam a formar-se muitas vezes, levando ao
encurtamento dos sarcómeros. A energia acumulada na cabeça das moléculas
de miosina permite a formação e movimento de pontes. Após a ocorrência do
movimento das pontes, o ATP tem que se ligar à cabeça de miosina. O ATP é
desdobrado em ADP e parte da energia é utilizada para libertar a ponte e fazer
com que a cabeça da molécula de miosina regresse à sua posição de repouso,
onde fica pronta para a formação de uma nova ponte. Alguma da energia do
ATP é armazenada na cabeça de miosina e utilizada para a formação e
movimento da ponte seguinte. A energia também é libertada sob a forma de
calor.
Fase de relaxamento
Os iões de cálcio são ativamente transportados para o retículo
sarcoplasmático.
Os complexos troponina-tropomiosina inibem a formação de pontes.
As fibras musculares alongam-se passivamente.
Intensidade do Estímulo e Contração Muscular
Em resposta a um estímulo apropriado, uma fibra muscular esquelética isolada produz
contrações de força igual em resposta a cada potencial de ação. A isto chama-se lei do
tudo ou nada da contração muscular e pode ser explicada com base na produção de
potenciais de ação na fibra muscular esquelética.
Quando se aplica na membrana da célula muscular um estímulo curto de força
crescente, acontecem os seguintes factos:
Um estímulo sublimiar não produz um potencial de ação, não ocorrendo
contração muscular.
Um estímulo limiar produz um potencial de ação, resultando na contração da
célula muscular.
Um estímulo supralimiar produz um potencial de ação da mesma amplitude
que o estímulo limiar e por isso produz uma contração idêntica.
As fibras musculares esqueléticas dispõem-se, no músculo esquelético, em unidades
motoras. Da mesma maneira que as fibras musculares isoladas, as unidades motoras
respondem segundo a lei do tudo ou nada, ou seja, todas as fibras musculares de uma
20
unidade motora se contraem para produzir uma força constante em resposta a um
estímulo limiar.
Os músculos no seu todo exibem características diferentes e mais complexas do que as
das fibras musculares isoladas ou das unidades motoras. Em vez de responder segundo
a lei do tudo ou nada, os músculos respondem a estímulos de uma maneira gradual, o
que significa que a força das contrações pode ir fraca, forte, consoante a força do
estímulo.
Um músculo compõe-se de muitas unidades motoras e os axónios das unidades
motoras combinam-se para formar um nervo. O músculo no seu todo contrai-se com
pouca ou com muita força consoante o número de unidades motoras recrutadas. Esta
relação chama-se somação de múltiplas unidades motoras, porque a força de
contração aumenta à medida que mais unidades motoras vão sendo estimuladas. À
medida que o estímulo aumenta pode acabar por se tornar num estímulo limiar. No
limiar, o estímulo já se tornou suficientemente forte para produzir um potencial de
ação no axónio de um neurónio motor e todas as fibras musculares dessa unidade
motora se contraem.
Estímulos progressivamente mais fortes, chamados estímulos submáximos, produzem
potenciais de ação em axónios de mais unidades motoras. Todas as unidades motoras
são ativadas por um estímulo máximo, valor em que um estímulo mais intenso, um
estímulo supramáximo, não tem efeito adicional. À medida que a força muscular
aumenta, entre o valor limiar e o valor máximo, são recrutadas unidades motoras, o
que significa que o número de unidades motoras que correspondem ao estímulo
aumenta e a força de contração produzida aumenta de forma contínua. No entanto,
cada unidade motora responde a potenciais de ação produzindo contrações de igual
amplitude.
21
Frequência do Estímulo e Contração Muscular
Uma fibra muscular isolada contrai-se em resposta a um potencial de ação. Emboras
um potencial de ação desencadeia a contração de uma fibra muscular, o potencial de
ação completa-se muito antes de se completar a fase de contração. Além disso, o
mecanismo contrátil numa fibra muscular não tem período refractário. Por isso, não é
necessário o relaxamento de uma fibra muscular antes de um segundo potencial de
ação poder estimular uma segunda contração. À medida que aumenta a frequência de
potenciais de ação numa fibra muscular esquelética, aumenta também a frequência
das contrações.
Quando as fibras musculares se relaxam parcialmente entra as contrações, fala-se em
tétano incompleto. Quando os potenciais de ação ocorrem com tal rapidez que não há
relaxamento muscular entre eles, designa-se a situação como tétano completo. A
tensão produzida por um músculo aumenta à medida que aumenta a frequência das
contrações. Esta tensão crescente chama-se somação de múltiplas ondas.
Outro exemplo de uma resposta variável é o fenómeno de ascensão em degraus que
se dá no músculo que esteve em repouso durante um período de tempo prolongado.
Se o músculo for estimulado com um estímulo máximo a baixa frequência, que
permite o relaxamento completo entre os músculos, a contração desencadeada pelo
segundo estímulo produz uma tensão ligeiramente maior do que o primeiro. A
contração desencadeada pelo terceiro estímulo produz uma tensão ligeiramente maior
do que o segundo. Só depois de alguns estímulos é que é igual a tensão produzida por
todas as contrações.
22
Tipos de Contração Muscular
Somação de múltiplas unidades motoras
Cada unidade motora responde segundo a lei do tudo-ou-nada.
Um músculo no seu todo é capaz de produzir uma força de contração
crescente, à medida que aumenta o número de unidades motoras estimuladas.
Somação de múltiplas ondas
A somação dá-se quando muitos potenciais de ação são produzidos numa fibra
muscular.
A contração dá-se em resposta ao primeiro potencial de ação, mas não há
tempo suficiente para que haja relaxamento muscular entre potenciais de ação.
Como cada potencial de ação produz a libertação de iões de cálcio do retículo
sarcoplasmático, os iões de cálcio mantêm-se elevados no sarcoplasma,
produzindo uma contração tetânica.
A tensão produzida em consequência da somação de múltiplas ondas é maior
do que a tensão produzida pela contração de uma única fibra muscular. O
aumento da tenção resulta da maior concentração de cálcio no sarcoplasma e
do estiramento dos componentes elásticos do músculo na fase precoce da
contração.
Tetania dos músculos
A tetania dos músculos resulta da somação de múltiplas ondas.
A tetania incompleta ocorre quando a frequência dos potenciais de ação é
suficientemente baixa para permitir o relaxamento parcial das fibras
musculares.
A tetania completa ocorre quando a frequência dos potenciais de ação é
suficientemente alta para não haver relaxamento das fibras musculares.
Fenómeno de ascensão em degraus
A tensão produzida aumenta durante as primeiras contrações em resposta a
um estímulo máximo, a baixa frequência, num músculo que ficou em repouso
durante um certo tempo.
A tenção crescente pode resultar da acumulação de pequenas quantidades de
cálcio no sarcoplasma durante as primeiras contrações, ou do aumento
crescente da atividade enzimática.
Contração isométrica
O músculo produz uma tensão crescente durante a contração.
O comprimento do músculo mantém-se contante durante a contração.
23
Este tipo é característico dos músculos posturais que mantêm uma tensão
constante sem mudarem o comprimento.
Contrações concêntricas
O músculo produz tensão crescente è medida que vai encurtando.
Contrações excêntricas
O músculo produz tensão, mas o comprimento muscular vai aumentando.
Tónus Muscular
É a tensão constante produzida pelos músculos do corpo durante longos períodos de
tempo. O tónus muscular é o responsável por manter eretos os membros inferiores e o
dorso, a cabeça levantada e o abdómen liso.
Comprimento vs. Tensão
A tensão ativa é a força aplicada a um objeto a ser levantado quando um músculo se
contrai.
O comprimento inicial de um músculo tem forte influência na quantidade de tensão
ativa que produz. Até um certo ponto, à medida que aumenta o comprimento de um
músculo, a sua tenção ativa aumenta também. Se o músculo for estirado para além do
comprimento ótimo, a tensão ativa que produz começa a declinar.
Se o músculo for estirado a ponto de os miofilamentos de actina e miosina não se
sobreporem ou se sobreporem em muito pouca extensão, o músculo produz muito
pouca tensão ativa quando estimulado.
Se o músculo não for estirado, os miofilamentos de miosina tocam cada uma das linhas
Z em cada sarcómero, podendo ocorrer aí muito pouca contração. Se o músculo está
estirado no seu comprimento ótimo, existe uma sobreposição ótima dos
miofilamentos. Quando o músculo é estimulado, a formação de pontes leva a uma
contração máxima.
Tensão passiva é a tensão aplicada a uma carga quando o músculo está estirado, mas
não estimulado. A tensão passiva existe porque o músculo e o seu tecido conjuntivo
possuem alguma elasticidade. A soma das tenções ativa e passiva é a tensão total.
24
Fadiga
Fadiga – Diminuição da capacidade de executar trabalho e a reduzida eficiência na
execução que se segue normalmente a período de atividade. A fadiga pode
desenvolver-se em três locais possíveis:
Sistema nervoso;
Músculos;
Junção neuromuscular.
Fadiga psicológica
É mediada pelo sistema nervoso central. Os músculos mantêm a sua capacidade
funcional, mas o indivíduo tem a sensação de que não é possível um trabalho muscular
suplementar.
Fadiga muscular
Resulta da depleção de ATP. Sem níveis adequados de ATP, as pontes não conseguem
funcionar normalmente. Em consequência, declina a tensão que o músculo é capaz de
produzir.
Fadiga sináptica
Dá-se na junção neuromuscular. Se a frequência de potenciais de ação for
suficientemente grande, a libertação de acetilcolina do terminal pré-sináptico é
superior à sua síntese. Deste modo, as vesículas sinápticas entram em depleção e a
acetilcolina libertada é insuficiente para estimular as fibras musculares.
25
Fontes de Energia
Creatina Fosfato
Em condições de repouso, a energia proveniente da respiração aeróbia é utilizada para
sintetizar a creatina fosfato. A creatina fosfato acumula-se nas células musculares e
destina-se a armazenar energia, que pode ser utilizada na síntese de ATP. Quando os
níveis de ATP começam a baixar, o ADP reage com a creatina fosfato para produzir ATP
e creatina.
ADP + Creatina fosfato Creatina + ATP
Glicólise
O metabolismo dos glícidos inicia-se com a glicólise que consiste numa série de
reações químicas no citosol da célula que desdobram uma molécula de glicose em
duas moléculas de ácido pirúvico.
A glicólise é dividida em quatro fases:
1) Entrada de ATP;
2) Clivagem do açúcar;
3) Produção de NADH;
4) Produção de ácido pirúvico.
26
1) Entrada de ATP
O primeiro passo da glicólise requer aporte de energia, na forma de ATP. Dá-se a
transferência de um grupo fosfato das moléculas de ATP para a molécula de glicose,
formando glicose 6-fosfato, por um processo chamado fosforilação.
Posteriormente os átomos de glicose 6-fosfato são reorganizados para formar frutose
6-fosfato, que é então convertida em frutose 1,6-difosfato pela adição de mais um
grupo fosfato de outra molécula de ATP.
2) Clivagem do açúcar
A frutose 1,6-difosfato (molécula com seis carbonos) é dividida em duas moléculas
com três carbonos:
Gliceraldeído 3-fosfato;
Dihidroxiacetona fosfato.
Esta segunda é restruturada e forma outra molécula de gliceraldeído 3-fosfato.
27
3) Produção de NADH
Cada molécula de gliceraldeído 3-fosfato é oxidada (perde dois eletrões) e forma
ácido 1,3-difosfoglicérico e a nicotinamida adenina dinucleótido (NAD+) é reduzida
(ganha dois eletrões) e forma NADH. O gliceraldeído 3-fosfatos também perde dois
iões de hidrogénio, um dos quais se liga ao NAD +.
NAD+ + 2e- + 2H + NADH + H+
28
4) Produção de ácido pirúvico
1- Transferência do grupo fosfato do ácido 1,3-difosfoglicérico para o ADP:
A enzima fosfogliceratoquinase transfere o grupo fosfato de alta energia do grupo
carboxilo do ácido 1,3-diofosfoglicérico para o ADP, formando ATP e ácido 3fosfoglicérico.
2- Conversão do ácido 3-fosfoglicérico em ácido 2-fosfoglicérico:
O ácido 3-fosfoglicérico sofre um rearranjo estrutural formando ácido 2-fosfoglicérico.
3- Desidratação do ácido 2-fosfoglicérico para ácido fosfoenolpirúvico:
Uma enzima, emolase, promove a remoção de uma molécula de água do ácido 2fofoglicérico, libertando ácido fosfoenolpirúvico.
4- Transferência do grupo fosfato do ácido fosfoenolpirúvico para o ADP:
O último passo na glicólise é a transferência do grupo fosfato do ácido
fosfoenolpirúvico para o ADP, formando, assim, ácido pirúvico e ATP.
29
30
Respiração Anaeróbia
A respiração anaeróbia consiste no desdobramento da glicose, na ausência de
oxigénio, para produzir duas moléculas de ácido láctico e duas moléculas de ATP. O
ATP assim produzido é uma fonte de energia durante atividades como o exercício
intenso, durante o qual é fornecida uma quantidade insuficiente de oxigénio aos
tecidos.
A respiração anaeróbica divide-se em duas fases:
1) Glicólise;
2) Formação de ácido láctico.
Formação de ácido láctico
Esta fase da respiração anaeróbia consiste na conversão de ácido pirúvico em ácido
láctico. As duas moléculas de NADH provenientes da glicólise cedem os 2H+ à molécula
de ácido pirúvico, formando ácido láctico.
31
Respiração Aeróbia
A respiração aeróbia consiste no desdobramento da glicose, na presença de oxigénio,
para produzir dióxido de carbono, água e 38 moléculas de ATP.
A respiração aeróbia divide-se em quatro fases:
1) Glicólise;
2) Formação de Acetil-CoA;
3) Ciclo de Krebs (ou ciclo do ácido cítrico);
4) Cadeia de transporte de eletrões.
Formação de Acetil-CoA
Nesta fase, o ácido pirúvico sai do citosol para uma mitocôndria, cujos
compartimentos interno e externo são separados pela membrana mitocondrial
interna. No compartimento interno, as enzimas removem um átomo de carbono e dois
átomos de oxigénio da molécula de ácido pirúvico com três carbonos, formando
dióxido de carbono e um grupo acetil com dois carbonos. Nesta reação é libertada
energia que é utilizada para reduzir o NAD+ a NADH. O grupo acetil combina-se com a
enzima A (CoA) para formal acetil-CoA.
Por cada duas moléculas de ácido pirúvico resultantes da glicólise são formadas duas
moléculas de acetil-CoA, duas moléculas de CO2 e duas de NADH.
32
Ciclo de Krebs
O ciclo de Krebs inicia-se com a produção de ácido cítrico, a partir da combinação de
acetil-CoA com uma molécula, com quatro carbonos, de ácido oxalacético. Ocorre
uma série de reações que formam novamente ácido oxalacético que, ao combinar-se
com outra molécula de acetil-CoA reinicia o ciclo.
Durante as reações que ocorrem no ciclo de Krebs, dão-se três acontecimentos
importantes:
1) Produção de ATP;
2) Produção de NADH e FADH2;
3) Produção de dióxido de carbono.
Produção de ATP
Por cada molécula de ácido cítrico é formada uma molécula de ATP.
Produção de NADH e FADH2
Por cada molécula de ácido cítrico são convertidas três moléculas de NAD+ em NADH e
uma molécula de flavina adenina dinucleótido (FAD) é convertida em FADH2.
Estas moléculas de NADH e FADH2 são transportadoras de eletrões, que entram na
cadeia de transporte de eletrões e são utilizadas para produzir ATP.
Produção de dióxido de carbono
Cada molécula de ácido cítrico com seis carbonos transforma-se, no final do ciclo de
Krebs, numa molécula de ácido oxalacético com quatro átomos de carbono.
Os dois átomos de carbono provenientes da molécula de ácido cítrico são utilizados
para formar duas moléculas de dióxido de carbono.
33
Cadeia de transporte de eletrões
Esta fase da respiração aeróbia consiste numa série de transportadores de eletrões na
membrana mitocondrial interna.
As moléculas de NADH e FADH2 cedem os seus eletrões para os transportadores e
libertam os iões H+ no compartimento interno na mitocôndria.
Os eletrões libertados pelo NADH e FADH2 movem-se ao longo da cadeia de
transportadores através de sucessivas reações de oxidação-redução. Três dos
transportadores de eletrões também funcionam como bombas de protões que
deslocam iões H+ do compartimento mitocondrial interno para o externo. Cada
bomba de protões recebe um eletrão, usa alguma da sua energia para fazer passar iões
hidrogénio para o compartimento externo e passa-o para o transportador seguinte.
O último transportador da série capta os eletrões e combina-os com oxigénio e iões
hidrogénio para formar água.
Uma vez que os iões hidrogénio libertados pelo NADH e FADH2 foram movidos para o
compartimento externo mitocondrial, a sua concentração nesse é superior á do
compartimento interno, difundindo-se novamente para o interno. Nesta difusão, os
iões hidrogénio passam através de canais formados pela enzima ATP sintease.
À medida que os iões se difundem de acordo com o gradiente de concentração, vão
perdendo energia que é utilizada para produzir ATP. Este processo denomina-se
modelo quimiosmótico.
O ATP é transportado para fora do compartimento interno por uma molécula
transportadora que troca ATP por ADP. Uma outra molécula transportadora conduz o
fosfato para o compartimento interno.
34
Resumo da produção de ATP
Por cada molécula de glicose, a respiração aeróbia produz um ganho líquido de 38
moléculas de ATP: 2 da glicólise, 2 do ciclo de Krebs e 34 na cadeia de transporte de
eletrões. Por cada molécula de NADH formada são produzidas três de ATP e por cada
molécula de FADH2 são produzidas duas na cadeia de transporte de eletrões.
Na respiração aeróbia são ainda produzidas 6 moléculas de dióxido de carbono. As
moléculas de água são reagentes nalgumas das reações químicas da respiração
aeróbia, e outras são produto. Das 12 moléculas de água produzidas são usadas 6, o
que se traduz num ganho líquido de 6 moléculas de água.
Equação da respiração aeróbia:
C6H12O6 + 6O2 + 38ADP + 38Pi 6CO2 + H2O + 38ATP
35
Fibras Lentas e Rápidas
Fibras Musculares de Contração Lenta ou Intensamente
Oxidativas
As fibras musculares de contração lenta, intensamente oxidativas ou fibras
musculares tipo I, contraem-se mais lentamente, são de menor diâmetro, têm uma
irrigação sanguínea mais bem desenvolvida, têm mais mitocôndrias e são mais
resistentes à fadiga do que as fibras musculares de contração rápida.
O metabolismo aeróbio é a fonte primária para a síntese de ATP nos músculos de
contração lenta e a capacidade destas para efetuar o metabolismo aeróbio é reforçada
por um abastecimento sanguíneo abundante e pela presença de numerosas
mitocôndrias.
Contêm mioglobina que fixa o oxigénio e atua como reservatório quando o sangue não
fornece quantidades adequadas.
Fibras Musculares de Contração Rápida ou Fracamente
Oxidativas
As fibras musculares de contração rápida, fracamente oxidativas ou fibras
musculares tipo II, respondem rapidamente à estimulação nervosa e contêm
moléculas de miosina que desdobram o ATP mais rapidamente do que as fibras
musculares de contração lenta. Têm uma irrigação sanguínea menos desenvolvida do
que os músculos de contração lenta.
Estas fibras têm grandes depósitos de glicogénio e estão bem adaptados à respiração
anaeróbia. Tendem a contrair-se rapidamente durante um período mais curto e
cansam-se relativamente depressa
36