OCB II
1–
1.1 - O sequenciamento do DNA é uma série de processos bioquímicos tem por finalidade determinar a ordem
dos nucleotídeos (adenina, guanina, citosina e timina) em uma amostra de DNA. Até meados da década de 70 era
muito complicado obter uma sequência de DNA, independente de ser fita simples ou fita dupla. No começo da
década de 80, foi desenvolvida uma técnica rápida de sequenciamento, por meio da quebra de uma cadeia de DNA
por diversos produtos químicos, sendo os fragmentos visualizados através do processo de eletroforese. Era
necessário marcar as moléculas com radiação, devido à baixa produção de material, não podendo ser detectada de
outra maneira.
Poucos anos após houve um novo avanço tecnológico foi conseguido por meio da introdução da técnica de
interrupção da sequência através da incorporação ao acaso de um nucleotídeo modificado, sendo chamada de
técnica de didesoxi ou dideoxi. Essa técnica, de imediato, tomou o lugar da anterior, possibilitando o
desenvolvimento de sequenciadores automáticos de DNA. O sequenciamento manual ainda existe, mas é muito mais
trabalhoso, caro e arriscado, pois utilizam-se substâncias radioativas.
Um dos métodos mais usados é a “técnica de desoxi”, também conhecida como terminadores de cadeia ou
Sanger (em homenagem a Fred Sanger, que propôs a técnica em questão).
A síntese de DNA se inicia por meio de um oligonucleotídeo, sendo continuada ao longo da seqüência do molde
pela enzima DNA polimerase, incorporada aos nucleotídeos e sintetizada a nova fita. Estão presentes nessa técnica
dois tipos de marcadores nucleotídeos: os “normais” (deoxi) – dATP, dGTP, dCTP e dTTP; e os dideoxinucleotídeos
(não possuem o grupo hidroxila no carbono 3’) – ddATP, ddGTP, ddCTP e ddTTP, sendo estes marcados com material
fluorescente.
A polimerização do DNA prossegue, sendo incluídos os nucleotídeos deoxi até o momento em que a DNA
polimerase, por acaso, insere um dideoxinucleotídeo, interrompendo assim a polimerização. Ao final de todo esse
processo são observados fragmentos de tamanhos distintos. Essa reação é transferida para um gel onde, após o
processo de eletroforese, é observada a migração dos fragmentos.
Quando o método utilizado é o manual, os dideoxinucleotídeos são marcados com radiação e não com
substâncias fluorescentes, sendo então necessária uma radiografia para a visualização da sequencia de DNA. Já no
sequenciamento automático, o seqüenciador identifica os fragmentos por meio da incidência de laser sobre os
dideoxinucleotídeos fluorescentes. Por fim, o seqüenciador mostra um gráfico onde cada nucleotídeo é referenciado
com cores distintas.
Depois que O DNA do Neandertal é sequenciado, é feito o exame de DNA comparativo entre as espécies que
consta
Inicialmente na separando o DNA do resto das células, por meio de detergentes e centrifugações. Em seguida,
aplicam-se pedaços de DNA sintético, marcados com corante, que se ligam aos trechos a serem examinados. Depois,
em um processo chamado eletroforese, os pedaços de DNA são separados por uma corrente elétrica de acordo com
seu tamanho. Finalmente, um equipamento a laser faz a leitura dos corantes e produz uma imagem computadorizada
que pode ser analisada e comparada pelos especialistas.
1.2 –
2
2.1 – As quatro rotas são: Respiração, Fermentação (respiração anaeróbia), fotossíntese e quimiossintese.
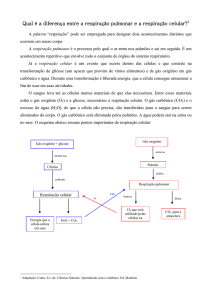
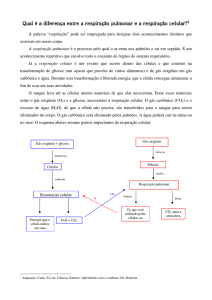
Respiração celular é o processo de conversão das ligações químicas de moléculas ricas em energia que poderão
ser usada nos processos vitais. Ela pode ser de dois tipos, Respiração Anaeróbia (sem utilização de oxigênio também
chamada de fermentação) e Respiração Aeróbia (com utilização de oxigênio). A respiração celular é o processo de
obtenção de energia mais utilizado pelos seres vivos. Na respiração, ocorre a libertação de dióxido de carbono e
energia e o consumo de oxigénio e glicose, ou outra substância orgânica. A organela responsável por essa respiração
é a mitocondria. O processo básico da respiração celular é a quebra da glicose ou Glicólise, que se pode
expressar pela seguinte equação química:
C6H12O6 + 6O2 → 6CO2 + 6H2O + energia
Nutrientes (energia química) + O2 → CO2 + H2O + Energia (alguma que se perde sob a forma de calor e
outra parte armazena-se sob a forma de ATP)
A respiração celular, que se processa segundo duas sequências básicas:
Glicose e Oxidação do piruvato através de um de dois processos: Respiração aeróbia ou Respiração anaeróbia
De acordo com o tipo de metabolismo, existem duas sequências possíveis para a oxidação do piruvato
proveniente da glicólise:
1. Neste primeiro processo, a oxidação do píruvato ocorre em uma respiração aeróbia, produzindo
em seguida Acetilcoenzima A, iniciando assim o Ciclo de Krebs, como é demonstrado a baixo: Piruvato -->
Acetil-Coa
Nesta etapa ocorre a entrada de NAD e CoA-SH.
O piruvato gerado na glicólise sofre desidrogenação e descarboxilação catalizado pelo complexo
piruvato-desidrogenase, durante essas reações é adicionada a coenzima A, desta forma a partir de cada
piruvato produz-se um acetil-CoA, iniciando logo em seguida o Ciclo de Krebs
Durante a glicólise pode ocorrer a falta de O2 gerando outras reações:
2. Nesta etapa a oxidação do piruvato ocorre em uma reação anaeróbia Oxidação Incompleta da
Glicose: Glicose --> Piruvato --> Ácido láctico ução não podem ser oxidados. Assim fica faltando NAD e FAD
para as reações de desidrogenação. Com isso, o ciclo de Krebs não pode ocorrer ficando totalmente
bloqueado, na falta de O2. Se houvesse uma forma de repor NAD e FAD sem o envolvimento da cadeia
respiratória o ciclo poderia continuar ocorrendo. Na oxidação da glicose na ausência de O2, o NADH
produzido não irá para a cadeia respiratória; da mesma forma, o piruvato não dara origem ao acetil-CoA.
Assim fica acumulado NADH e piruvato. Para que a glicólise mantenha-se, o NADH acumulado transfere
seus elétrons e P+ para o piruvato, originando ácido láctico e renegerando o NAD. Isto representa uma via
alternativa de oxidação do NADH. Na oxidação incompleta o rendimento de ATP cai para apenas dois.
[editar] Respiração aeróbia
A respiração aeróbia requer oxigênio. Na glicólise, é formado o piruvato (também chamado de ácido
pirúvico) bem como 2 ATP. Cada piruvato que entra na mitocôndria e é oxidado a um composto com 2
carbonos (acetato) que depois é combinado com a Coenzima-A, com a produção de NADH e libertação de
CO2. De seguida, inicia-se o Ciclo de Krebs. Neste processo, o grupo acetil é combinado com compostos com
4 carbonos formando o citrato (6C). Por cada ciclo que ocorre liberta-se 2CO2, NADH e FADH2. No ciclo de
Krebs obtém-se 2 ATPs. Numa última fase - cadeia transportadora de elétrons (ou fosforilação oxidativa) os
elétrons removidos da glicose são transportados ao longo de uma cadeia transportadora, criando um
gradiente protónico que permite a fosforilação do ADP. O aceptor final de elétrons é o O2, que, depois de se
combinar com os elétrons e o hidrogênio, forma água. Nesta fase da respiração aeróbia a célula ganha 32
moléculas de ATP. Isso faz um total ganho de 30 ATP durante a respiração celular em que intervém o
oxigênio.
A respiração anaeróbia envolve um receptor de elétrons diferente do oxigênio e existem vários tipos
de bactérias capazes de usar uma grande variedade de compostos como receptores de elétrons na
respiração: compostos nitrogenados, tais como nitratos e nitritos, compostos de enxofre, tais como
sulfatos, sulfitos, dióxido de enxofre e mesmo enxofre elementar, dióxido de carbono, compostos de ferro,
de manganês, de cobalto e até de urânio.
No entanto, para todos estes , a respiração anaeróbia só ocorre em ambientes onde o oxigénio é
escasso, como nos sedimentos marinhos e lacustres ou próximo de nascentes hidrotermais submarinas.
Uma das sequências alternativas à respiração anaeróbia é a fermentação, um processo em que o
piruvato é apenas parcialmente oxidado, não se segue o ciclo de Krebs e não há produção de ATP numa
cadeia de transporte de elétrons. No entanto, a fermentação é útil para a célula porque regenera o
dinucleótido de nicotinamida e adenina (NAD), que é consumido durante a glicólise.
Os diferentes tipos da fermentação produzem vários compostos diferentes, como o etanol (o álcool
das bebidas alcoólicas, produzido por vários tipos de leveduras e bactérias) ou o ácido láctico do iogurte.
Outras moléculas, como NO2, SO2 são os aceptores finais na cadeia de transporte de elétrons.
A fotossíntese é o processo através do qual as plantas, seres autotróficos (seres que produzem seu próprio
alimento) e alguns outros organismos transformam energia luminosa em energia química processando o dióxido de
carbono (CO2), água (H2O) e minerais em compostos orgânicos e produzindo oxigênio gasoso (O2).
A equação simplificada do processo é a formação de glicose e libertação de oxigênio: 6H2O + 6CO2 → 6O2
+C6H12O6.
Este é um processo do anabolismo, em que a planta acumula energia a partir da luz para uso no seu
metabolismo, formando adenosina tri-fosfato, o ATP, a moeda energética dos organismos vivos.
A fotossíntese inicia a maior parte das cadeias alimentares na Terra. Sem ela, os animais e muitos outros seres
heterotróficos seriam incapazes de sobreviver porque a base da sua alimentação estará sempre nas substâncias
orgânicas proporcionadas pelas plantas verdes.
Na fase fotoquímica, fase luminosa ou fase clara (fase dependente da luz solar) é a primeira fase do processo
fotossintético. A energia luminosa é captada por meio de pigmentos fotossintetisantes, capazes de conduzi-la até o
centro de reação. Tal centro é composto por um par de clorofilas 'a' também denominado P700 porque absorve a onda
luminosa com 700 nanometros de comprimento. Os elétrons excitados da P700 saem da molécula e são transferidos
para uma primeira substância aceptora de elétrons, a ferredoxina. Esta logo os tranfere para outra substância, e
assim por diante, formando uma cadeia de transporte de elétrons. Tais substâncias aceptoras estão presente na
membrana do tilacóide. Nessa transferência entre os aceptores, os elétrons vão liberando energia gradativamente e
esta é aproveitada para transportar hidrogênio iônico de fora para dentro do tilacóide, reduzindo o pH do interior
deste. A redução do pH ativa o complexo protêico "ATP sintetase". O fluxo de hidrogênios iônicos através do
complexo gira, em seu interior, uma espécie de "turbina proteica", que promove a fosforilação de moléculas de
adenosina difosfato dando origem à adenosina trifosfato. Ao chegarem ao último aceptor, os elétrons têm nível
energético suficientemente baixo e retornam ao par de clorofilas 'a', fala-se em fotofosforilação cíclica.
Porém, existe outra forma de fosforilação, a fotofosforilação acíclica onde os elétrons das moléculas de
clorofila 'a' (P700), excitados pela luz, são captados pela ferredoxina, mas ao em vez de passarem pela cadeia
transportadora são captados pelo NADP (nicotinamida adenina dinucleotídeo Fosfato) e não retornam para o P700.
Este fica temporariamente deficiente de elétrons. Esses elétrons são repostos por outros provinientes de outro
fotossistema onde o par de clorofilas 'a', dessa vez P680, excitado pela energia luminosa, libera elétrons que são
captados por uma primeira substância aceptora: a plastoquinona. Em seguida passa aos citocromos e plastocianina
até serem captados pelo P700, que se recompõe. Este processo de transporte também promove a síntese do ATP. Já
o P680 fica deficiente de elétrons. Esses elétrons serão repostos pela fotólise da água. A quebra da molécula da água
por radiação (fotólise da água) produz iôns de hidrogênios e hidróxidos. Os elétrons dos iôns hidróxidos são utilizados
para recompor o P680 e os iôns hidrogênio são aceptados pelo NADP, com isso ocorre a formação de água oxigenada
(H2O2) oriunda da reação de síntese entre as hidroxilas. A água oxigenada é decomposta pela célula em água e O2
sendo este último liberado do processo como resíduo. Com a repetição do processo forma-se o aporte energético e
de NADPHs necessários para a fase escura.
A fase química ou "fase escura", onde se observa um ciclo descoberto pelos cientistas Melvin Calvin, Andrew
Benson e James Bassham. Nessa fase chamada de ciclo de Calvin ou ciclo das pentoses, que ocorre no estroma do
cloroplasto, o tilacóide fornece ATP e NADPH2 ao estroma do cloroplasto, onde se encontra a pentose (ribulose
fosfato), essa pentose ativada por um fosfato, fixa o carbono que provém do dióxido de carbono do ar sob a ação
catalisadora da "rubisco" (ribulose bifosfato carboxilase-oxidase) e em seguida é hidrogenada pelo NAPH2 formando
o aldeído que dará origem à glicose. Para a síntese de uma molécula de glicose são fixadas seis de dióxido de
carbono, permitindo que o processo recicle a ribulose fosfato. devolvendo-a ao estroma. Desta fase resulta a
formação de compostos orgânicos como a glicose, necessária à atividade da planta. Esta fase é denominada fase
escura, no entanto é um termo utilizado de forma inadequada pois para a "rubisco" entrar em atividade
determinando a fixação do CO2 atmosférico para a formação de moléculas de glicose, ela precisa estar num estado
reduzido, e para isso acontecer é necessário que a luz esteja presente.
Este processo consiste na sintese de substancias organicas a partir da energia libertada em certas reações
quimicas inorganicas.
A quimiossintese é um processo analogo à fotossintese, diferindo fundamentalmente na fonte de energia que
gera um fluxo de elétrons e prótons, permitindo a fosforilação do ADP a ATP e a redução de um transportador de
hidrogénios.
As bacterias quimiossintetizantes conseguem desenvolver-se em ambientes desprovidos de luz e materia
organica, pois a energia necessaria ao seu desenvolvimentos é obtida atraves de oxidaçoes inorganicas. Apenas
necessitam de oxigenio, para a oxidaçao, e de dioxido de carbono e água como materia prima para a sintese de
glicose.
As nitrossomas e as nitrobacterias sao bacterias quimiossintetizantes que participam na reciclagem do azoto no
nosso planeta. As nitrossomonas obtem energia atraves da oxidaçao de amônia presente no solo, transformando-a
em nitritos. as nitrobacterias, por sua vez, utilizam o nitrito, oxidando-o a nitrato.
3 – A conjugaçao, ou seja, união parcial transitória de dois indivíduos, na qual se trocam mutuamente núcleos
haplóides de modo que, depois de realizada a separação, os núcleos dos ex-conjugantes possuem uma nova
guarnição cromossômica combinada. A conjugação encontra-se somente nos ciliados, os protozoários mais altamente
diferenciados e mais ricos em diferenciação citoplasmática. O ciclo é diplobionte.
Dois ciliados, na maior parte das vezes com a mesma forma, encontra-se um ao outro pela região oral; aí se
forma uma ponte citoplasmática.
Após a união pelo citóstoma, o macronúcleo de cada um se desorganiza e desaparece, enquanto o micronúcleo
sofre duas divisões sucessivas, ou seja, meiose.
- Dos 4 núcleos resultantes, 3 degeneram e o último se divide mais uma vez; os 2 núcleos finais são
chamados de PRONÚCLEOS: femininos e masculinos.
- Os pronúcleos masculinos de um do outro migram para a célula de seu parceiro e se unem com os
pronúcleos femininos que haviam permanecido no lugar. O núcleo resultante da fusão em cada célula é um zigoto
(syncarion) onde o número diplóide de cromossomos é restabelecido.
- As células se separam e cada uma delas se divide 3 vezes seguidas, produzindo 8 núcleos filhos.
- As duas primeiras divisões são meióticas
- Dos 8 núcleos, 4 aumentam de tamanho até se transformar em outros tantos macronúcleos
- Um fica sendo o micronúcleo e os demais desaparecem.
- Seguem-se duas divisões normais do micronúcleo e de toda a célula (porém sem multiplicação dos
macronúcleos), dando como resultado final a produção de 4 ciliados a partir de um dos 2 ex-conjugantes, com um
macronúcleo e um micronúcleo cada.
Os parceiros da conjugação são hermafroditas, pois fornecem núcleos gaméticos de dois tipos, que se
comportam diferentemente. Os paramécios, porém, só conjugam quando pertencem a diferentes, mas compatíveis,
tipos de emparelhamento (Matingtypes),. Antes da conjugação há emissão de sinais químicos por alguns ciliados, mas
em outros, como Paramecium, a substância química permanece na superfície celular a qual expressa a resposta
quando os ciliados fazem contacto físico.
4–
4.1 – o domínio Eukarya inclui todos os organismos eucariontes; o domínio Eubacteria ou simplesmente
Bacteria inclui todas as bactérias (procariontes); o domínio Archaea (anteriormente designado "Archaebacteria")
inclui procariontes com características filogenéticas (relações evolucionárias obtidas por análise genómica) diferentes
da bactérias.
4.2 – Eukarya
4.3 – Carioteca, organelas membranosas
4.4 - Flora intestinal é um nome popular dado a microbiótica intestinal que consiste em um conjunto de
bactérias (Eubacteria) que naturalmente habitam o intestino, ajudando na digestão de alimentos e controlando (por
competição) o crescimento de microrganismos patogênicos (outras bactérias, fungos, protozoários, etc).
4.5 – Todos sao dotados de celulas.
5–
5.1 - O ciclo foi primeiro elucidado por Calvin e colaboradores em 1946 e por esta razão, também é conhecido
como ciclo de Calvin. Ele pode ser dividido em quatro fases distintas: fase de carboxilação, fase de redução, fase de
regeneração e fase de síntese dos produtos. A fase de carboxilação consiste na reação de CO2 com a ribulose
bisfosfato, catalisada pela ribulose-1,5-bisfosfato carboxilase (RuBisCO), seguida por uma clivagem molecular,
formando o ácido fosfoglicérico. A fase de redução consiste na redução do ácido glicérico, formado na etapa anterior,
em triose fosfato. A fase de regeneração consiste na regeneração da ribulose bisfosfato através de reações de
interconversão de açúcares. A fase de síntese de produtos consiste na produção de outros compostos, tais como,
polissacarídeos, aminoácidos e ácidos graxos. A síntese desses compostos é influenciada pelas condições fisiológicas.
O ciclo de Calvin também é conhecido como a rota C3 de fixação do carbono, uma vez que o produto formado
é um composto de 3 carbonos (ácido fosfoglicérico). Entretanto, esta não é a única rota de fixação do CO2. Na
maioria das plantas e gramíneas tropicais, tais como, a cana-de-açúcar e a cevada, a fixação do CO2 resulta em
compostos de 4 carbonos como o oxaloacetato, o malato e o aspartato. A fixação ocorre pela carboxilação do
fosfoenolpiruvato a oxaloacetato catalisada pela fosfoenolpiruvato carboxilase. Por essa razão, essa rota é chamada
de C4. Existe ainda o metabolismo ácido das crassuláceas (CAM), cujo nome se deve ao fato de ser primeiro
encontrado nas Crassulaceae. Esta rota de fixação do CO2 é muito comum nas famílias das angiospermas: Agavaceae,
Bromeliaceae, Cactaceae, Euphorbiaceae, Liliaceae, Orchidaceae, etc. Como nas plantas de metabolismo C4, o
primeiro metabólito a ser sintetizado pela fixação do CO2 é o oxaloacetato. Este CO2 é posteriormente liberado pela
descarboxilação do malato e refixado no ciclo de Calvin pela RuBisCO. Entretanto os metabolismos CAM e C4 diferem
entre si pelo local e tempo de ocorrência. Nos vegetais que apresentam metabolismo C4, a fixação do CO2 ocorre nas
células fotossintéticas presentes no mesófilo da folha. O carbono fixado na forma de malato migra para as células
envolventes da bainha onde ocorre então a liberação e refixação do CO2 através do ciclo de Calvin. Nas plantas do
metabolismo CAM o período de fixação via fosfoenolpiruvato carboxilase e RuBisCO estão separados pelo tempo.
Nessas plantas, a fixação ocorre durante a noite quando os estômatos estão abertos via carboxilação do
fosfoenolpiruvato e acúmulo do malato, assim formado, nos vacúolos. Durante o dia, os estômatos se fecham para
minimizar a perda de água, e o malato é transportado para o citossol onde é descarboxilado e o CO2 é refixado pela
RuBisCO.
As vantagens dos metabolismos CAM e C4 sobre o C3 são: alta taxa fotossintética (dificilmente atinge-se a
saturação da fotossíntese), ausência de fotorrespiração, alta eficiência na utilização da água, alta tolerância salina e
baixo ponto de compensação para o CO2. A desvantagem é o alto custo energético e o conseqüente menor
rendimento quântico de fixação de CO2. Além disso, com exceção do abacaxi, as plantas de metabolismo CAM, ao
contrário das de metabolismo C3, não são muito produtivas em termos de biomassa. Os vegetais de metabolismo C4
são altamente produtivos.
As macroalgas além de possuírem o metabolismo C3, cujo produto principal nas Phaeophyta é o manitol, são
capazes de fixar CO2 ou HCO3- independente de luz, pela carboxilação do fosfoenolpiruvato a malato. Entretanto, ao
contrário das plantas de metabolismos C4 e CAM, onde o CO2 fixado é posteriormente liberado para o ciclo de Calvin,
o oxaloacetato formado pela fixação do CO2, via fosfoenolpiruvato, é utilizado para as reações de biossíntese ou
como intermediário do ciclo do ácido tricarboxílico, havendo portanto, fixação líquida de CO2. Esta capacidade de
fixar CO2 independente de luz é ecologicamente significante, pois permite o crescimento das macroalgas (em
particular a Phaeophyta, onde este tipo de fixação é expressiva) em ambientes de irradiância limitada. As outras rotas
metabólitas acopladas com a fase clara da fotossíntese são: a fixação do N2, a biossíntese de ácidos graxos, a reação
de Mehler e a fotorrespiração.
A fotorrespiração é uma ineficiência do ciclo de fixação de CO2. A Enzima RuBisCO não é específica para o CO2,
ela também aceita como substrato o O2, de sorte que, ao invés de ser produzido 2 moléculas de ácido glicérico após
a clivagem molecular, produz-se uma molécula de ácido glicérico e uma molécula de ácido glicólico, que na célula
está na forma aniônica glicolato. O glicolato é exportado aos peroxomas onde é oxidado à glioxalato e então aminado
para formar o aminoácido glicina (Gli). A glicina por sua vez, é exportada para a mitocôndria onde 2 moléculas de
glicina são utilizadas para gerar uma molécula de serina (Ser) e uma molécula de CO2. Daí o nome fotorrespiração,
pois há a participação da luz (foto), o consumo de O2 e a liberação de CO2 (respiração). A fotorrespiração é
favorecida em ambientes de alta concentração de O2 e baixa de CO2 (condição atmosférica) e em altas temperaturas
(Climas tropicais e equatoriais).
5.2 – Maior fixação de carbono, implica em uma maior produção de biomassa que será repassado ao longo da
cadeia alimentar, juntamente com uma maior liberação de oxigênio.