GENÉTICA DE TRANSMISSÃO:
1ª E 2ª LEI DE MENDEL
13
GENÉTICA DE TRANSMISSÃO
1. LEI DA SEGREGAÇÃO DOS FATORES, 1ª LEI DE MENDEL, OU LEI DA PUREZA
DOS GAMETAS
Figura 1.1: vagem e flor da ervilha, primeiro vegetal utilizado nos estudos da genética.
Fonte: Snustad, D.Peter; Simmons, Michael J.,2008.
INTRODUÇÃO
Apesar de há milhares de anos se tentar explicar como as características
genéticas são transmitidas, só no fim século XIX, obteve-se os primeiros resultados
significativos a respeito de como essas características são passadas de pais para
filhos, surgindo dessa forma o que hoje chamamos de genética de transmissão.
Nesse capítulo, falaremos sobre os trabalhos de Mendel, sua metodologia e seus
postulados, que serviram de ponto de partida para o entendimento dessa ciência.
1.1. QUEM FOI MENDEL!
Gregor Johann Mendel nasceu em 1822, no vilarejo de Heinzendorf, antiga
Áustria e hoje República Tcheca, filho de pais agricultores pobres, deve uma
criação rural o que facilitou o seu conhecimento sobre os processos de cultivo de
plantas, de criação de animais e seu amor pela natureza. A falta de condições
14
financeiras para estudar fez com que Mendel aos 21 anos optasse por um
monastério católico na cidade de Brünn (hoje, Brno) para dar continuidade aos
seus estudos. Quatro anos mais tarde ordenou-se padre (1847), adotando o nome
Gregor pela igreja. Durante, esses quatro anos, Mendel aprendeu ciências agrárias
e técnicas de polinização. Entre 1851 e 1853 foi liberado por seus superiores do
mosteiro, para cursar na Universidade de Viena o curso de Física, porém assistiu
cursos adicionais de química, zoologia, botânica, fisiologia vegetal, paleontologia e
de Matemática, do qual, já tinha sido professor numa escola local, próxima ao
mosteiro. Também se dedicou ao estudo de técnicas de hibridização em plantas e
em especial em ervilhas. Em 1853, quando retornou ao Mosteiro, voltou as suas
atividades de monge-professor, agora de física e ciências naturais e começou em
1857, seus experimentos genéticos com ervilha, concluindo-os em 1864. Em 1865
apresentou os seus resultados na Natural History Society local e no ano seguinte,
publicou um relato detalhado nas publicações da sociedade. Mendel, porém, ficou
na obscuridade por 35 anos, não se sabe se por falta de entendimento do que
publicou na época, ou por que os interesses dos cientistas da época estavam
voltados para outras questões, como por exemplo, a evolução.
Figura 1.2: Gregor Johann Mendel (1822 – 1884). Fonte Klug et. al.,2010
Em 1900, três botânicos, Hugo de Vries na Holanda, Carl Correns na
Alemanha e Eric Von TschermaK-Seysenegg na Áustria de forma independente,
trabalhando com hibridização em outros vegetais descobriram ao pesquisarem a
15
literatura científica, para corroborar com suas teorias sobre hereditariedade, que
Mendel há 35 anos tinha chegado às mesmas conclusões.
1.2. PORQUE MENDEL OBTEVE SUCESSO?
Várias foram as causas do sucesso de Mendel, a escolha do material, o seu
alto grau de organização científica na escolha da metodologia e seus
conhecimentos estatísticos, como conhecedor e professor de matemática por um
determinado tempo.
O material biológico escolhido por Mendel foram variedades de plantas da
espécie Pisum sativum (34 variedades, cedidas por horticultores da região),
também
chamadas ervilhas-de-jardim ou ervilhas-de-cheiro
(figura 1.3),
dicotiledôneas de fácil cultivo, podem ser plantadas em jardins experimentais, ou
em vasos, em uma estufa. Por apresentarem suas pétalas em forma de quilhas
(fechadas) são impedidas de realizarem fecundação cruzada de forma natural,
sendo assim, hermafroditas e autofecundantes, o que facilita a obtenção de
linhagens puras para uma determinada característica. Porém, é acessível para
cruzamentos experimentais de hibridização, originando híbridos férteis. Seu ciclo
de vida é curto, podendo originar várias gerações, em pouco tempo. Apresentam, a
cada geração, um grande número de descendentes, o que facilita o estudo
estatístico dos dados colhidos. Apresentam linhagens com características
individuais bem contrastantes, tais como cor dos cotilédones da semente que em
uma linhagem é verde e em outra linhagem amarela; altura do pé de ervilha que
poderia ser alto (em torno de 2,0 m de altura) ou baixo (em torno de 0,5 m de
altura) e assim sucessivamente para as sete características estudas por ele(figura
1.4).
16
Figura 1.3: Flor hermafrodita da ervilha, com identificação das pétalas fechadas em
forma de quilha, isolando do meio esterno as estruturas reprodutora masculina e
feminina da flor. Fonte: Amabis, José M.,Martho, Gilberto R., 2006.
Figura 1.4: As sete características estudadas por Mendel. Fonte: Griffiths et. al., 2009
Com relação à metodologia, Mendel preocupou-se em trabalhar na
observação de uma característica por vez, anotando seus resultados e
comparando-os com os das outras seis características estudadas, além de obter,
amostras significativas para a análise estatística dos dados, decorrência do grande
número de descendentes obtidos em cada geração.
17
1.3. UM EXEMPLO DO EXPERIMENTO DE MENDEL
Um dos experimentos realizado por Mendel foi o cruzamento entre uma
variedade de ervilhas que durante várias gerações só produzia, por
autofecundação, sementes com cotilédones amarelos, com uma variedade que só
produzia cotilédones verdes, sendo assim denominadas sementes puras, para
essa característica analisada. Em cada experimento Mendel retirava as anteras,
estruturas vegetais onde se produz o grão de pólen, de algumas das plantas com
cotilédones amarelos, ficando essas variedades só com a estrutura reprodutora
feminina. Repetia o mesmo processo com as variedades de cotilédones verdes.
Como resultado em cada uma das variedades estudadas existiriam plantas
somente com estruturas reprodutoras femininas e outras hermafroditas. Após esse
processo de castração, Mendel esperava o amadurecimento reprodutivo das
plantas femininas e realizava a fertilização cruzada, retirando o pólen das anteras
das plantas de cotilédones verdes e colocando nas plantas femininas de
cotilédones amarelos, do mesmo jeito, retirava o pólen das de cotilédones
amarelos e fertilizava as femininas de cotilédones verdes (figura 1.5), realizando
cruzamentos recíprocos, nos dois casos, os resultados obtidos foram os mesmos:
os descendentes, todos nasceram com cotilédones amarelos. Essas plantas
resultantes da fertilização cruzada foram denominadas híbridas.
Figura 1.5: Polinização cruzada e autofecundação os dois tipos de cruzamentos usados
por Mendel. Fonte: Griffiths et. al., 2009
18
O passo seguinte foi deixar as plantas híbridas autofecundarem para se
observar a descendência. A característica cotilédone verde que havia desaparecido
na geração anterior (híbrida) voltou a aparecer numa proporção aproximada de
uma semente de cotilédone verde para cada três com cotilédone amarelo (fig. 1.6).
Mendel chamou o 1º cruzamento entre linhagens puras de geração
parental, hoje chamada simplesmente geração P, a prole desse 1º cruzamento,
descendência híbrida, geração filial 1 ou F1 e os descendentes da autofecundação
de F1, geração filial 2, ou F2 e assim sucessivamente.
Figura 1.6: Representação esquemática das gerações P, F1 e F2 do cruzamento monohíbrido para a característica cor do cotilédone da semente Fonte: Amabis, José
M.,Martho, Gilberto R., 2006.
Completando o experimento, Mendel pegou as sementes com cotilédones
verde da geração F2 deixou germinar e autofecundar, obtendo em F3, sementes
com cotilédones verdes, já as sementes com cotilédones amarelos da geração F2, ao
19
germinar e autofecundar gerou em 1/3 dos pés de ervilha somente sementes com
cotilédones amarelos e em outros 2/3, sementes com cotilédones amarelos e
verdes.
Mendel repetiu esse experimento, denominado cruzamento mono-híbrido
(pois só leva em consideração uma única característica ou caráter por vez), com as
outras seis características contrastantes da ervilha-de-jardim, obtendo sempre o
mesmo resultado. Em F1, no híbrido, só uma das características aparecia; em F2, a
proporção era sempre de aproximadamente 3 com a mesma característica de F1
para 1 com a característica que desaparecia em F1 (fig.1.7).
Mendel também vez o cruzamento recíproco entre as plantas híbridas, F1
com as plantas parentais de ervilhas de cotilédones verdes, obtendo na prole a
proporção de 1 ervilha de cotilédone amarela, para uma ervilha de cotilédone
verde. Hoje esse cruzamento é conhecido como cruzamento teste (fig. 1.8).
20
Plantas F1
(híbrida)
Autofecundação de
F1(híbridas)
Forma das sementes
Lisa X Rugosa
Sementes lisas
Lisas X Lisas
2
Cor dos cotilédones
Amarelo X Verde
Sementes com
cotilédones
amarelo
Amarelo X Amarelo
3
Cor da flor
Violeta X branca
Flores violeta
Violeta X Violeta
4
Textura das vagens
Inflada X comprimida
Vagens infladas
Inflada X Inflada
5
Cor das vagens
Verdes X Amarelas
Vagens verdes
Verde X Verde
6
Posição das flores
Axilar X Terminal
Flores axilares
Axilar X Axilar
Nº de
características
Geração
Parental(pura)
( P)
1
7
Altura do caule
Alto X baixo
Caule alto
Alto X Alto
Plantas F2
Razão entre
os tipos F2
5474 lisas e 1850
rugosas=
7324(total)
6022com
cotilédones
amarelo e 2001
com cotilédones
verdes= 8023
(total)
705 flores
violetas e 224
flores brancas=
929 (total)
882 vagens
infladas e 299
vagens
comprimidas
= 1181 (total)
428 vagens
verdes e 152
vagens amarelas=
580 (total)
651 flores
axilares e 207
flores terminais=
858 (total)
2,96: 1
3,01: 1
3,15: 1
2,95:1
2,82: 1
3,14:1
787 caule alto e
277 caule baixo =
1064(total)
2,84:1
Figura 1.7: Resultados dos cruzamentos Mendelianos nos quais os genitores diferem em
uma característica. Adaptada Fonte: Amabis, José M.,Martho, Gilberto R., 2006.
21
Figura 1.8: Esquema da autofecundação de F1, híbrida, aparecendo na prole a
proporção de 3: 1(esquerda) e o cruzamento teste entre F1, híbrida, e a planta de
cotilédones verdes dando a proporção de 1:1 (direita). Fonte: Griffiths et. al., 2009
1. 4. CONCLUSÕES DE MENDEL
Após análise estatística dos dados Mendel chegou às seguintes conclusões:
1º) Existem fatores unitários particulados, que funcionam como unidades
básicas da hereditariedade (hoje chamamos genes) responsáveis por
cada uma das características estudadas e são transmitidas de geração a
geração de forma inalterada através dos gametas; (Exemplo: Um fator
determina a cor do cotilédone verde e outro fator determina a cor do
cotilédone amarelo)
22
2º) Cada planta ou organismo individual possuem um par desses fatores,
determinando a característica; (Exemplo cor do cotilédone)
3º) Na hora de formar os gametas só um dos fatores será encontrado nos
gametas;
4º) Na fertilização após o encontro do gameta masculino (presente no
pólen) com o gameta feminino (presente no pistilo) haverá uma
reconstituição do par de fatores.
5º) Cada planta pura, para uma dada característica possuirá um par de
fatores iguais;
6º) A planta híbrida apresentará um fator de cada tipo na formação do par;
7º) O fator que não aparece na geração F1, e só volta a reaparecer na geração
F2 é dito recessivo, e o fator que determina a característica em F 1 é dito
dominante.
1.5.
A
CORRELAÇÃO
ENTRE
OS
POSTULADOS
DE
MENDEL
E
O
COMPORTAMENTO DOS CROMOSSOMOS NA MEIOSE
As redescobertas dos trabalhos de Mendel em 1900 por Hugo de Vries na
Holanda, Carl Correns na Alemanha e Eric Von TschermaK-Seysenegg na Aústria, e
sua aceitação pelos cientistas originaram o levantamento de outras questões, tais
como: onde se localizam,nas células, os fatores hereditários ? Qual é o mecanismo
responsável por sua segregação durante a formação dos gametas.
Em 1902,Walter S. Sutton, trabalhando com a formação de gametas em
gafanhoto e Theodor Bovari trabalhando de forma independente, com meiose,
observaram uma grande semelhança entre o comportamento dos cromossomos na
meiose e a segregação dos fatores hereditários (genes) de Mendel, Sutton e Bovari
propuseram assim a hipótese de que os fatores hereditários de Mendel estavam
localizados em cromossomos homólogos, de tal maneira que sua separação na
meiose levaria à segregação dos fatores.(fig. 1.9)
23
Figura 1.9: Representação esquemática da idéia originalmente proposta por Walter S.
Sutton e Bovari, em 1902, de que a segregação de um par de alelos resulta da separação
dos cromossomos homólogos na meiose. A hipótese foi confirmada e passou a
constituir um dos fundamentos da Genética. Fonte: Amabis, José M.,Martho, Gilberto R.,
2006.
24
1.6. A TERMINOLOGIA GENÉTICA ATUAL
Hoje sabemos que os fatores hereditários de Mendel são os genes, que
podem ser representado simbolicamente de várias formas, adotaremos
inicialmente, de forma simples, a letra da característica recessiva, na forma
minúscula, e em itálico, como símbolo representativo do traço recessivo
(por
exemplo: Na característica cor do cotilédone, v = fator ou gene que determina a
característica ou traço cotilédone verde). O traço dominante da mesma
característica (cotilédone amarelo) é representado pela mesma letra, também em
itálico, só que maiúscula (V).
Os fatores hereditários alternativos que determinam traços diferentes
dentro de uma mesma característica (V e v) são denominados alelos. Logo cada
planta apresenta dois alelos para uma dada característica (VV ou Vv ou vv). Esses
pares de alelos representam o genótipo ou constituição gênica da planta para a
característica estudada. Os traços cotilédones amarelos ou cotilédones verdes,
variações dentro de uma mesma característica e expressão física dos fatores
hereditários, ou genes, é denominado fenótipo.
O fenótipo cotilédone amarelo dominante em relação ao fenótipo cotilédone
verde, apresenta dois genótipos diferentes, assim representados: VV encontrado
em plantas puras ou hoje chamadas homozigotas para essa característica, pois só
produzem um tipo de gameta V; e o genótipo Vv, encontrado em plantas híbridas
ou heterozigotas, capaz de produzirem dois tipos de gametas V ou v. O fenótipo
recessivo, cotilédone verde apresenta um único genótipo vv originando um único
tipo de gameta v.(fig. 1.10)
25
Figura 1.10: Esquema de um cruzamento mono-híbrido de ervilha-de-jardim,
representando a característica cor do cotilédone Amarelo(V) e verde(v). Adaptada de
Klug et. al., 2010.
26
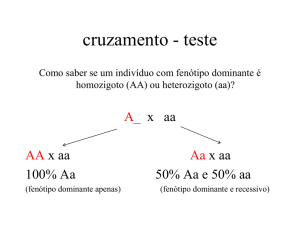
1.7. CRUZAMENTO TESTE
O cruzamento teste, feito por Mendel, cruza um fenótipo dominante com um
recessivo e serve para identificar se o fenótipo dominante tem genótipo
homozigoto ou heterozigoto, quando obtemos toda a prole do cruzamento com
fenótipo dominante, podemos chegar a conclusão que o fenótipo dominante é
homozigoto, já quando obtemos aproximadamente metade da prole com o fenótipo
dominante e a outra metade recessiva o genótipo do fenótipo dominante é
heterozigoto.
1.8. OS QUADROS DE PUNNETT
Reginald C. Punnett construiu um diagrama que facilita visualização de um
cruzamento, nesse diagrama representamos os gametas masculinos em uma
coluna ou linha. Os gametas femininos, são representados dependendo dos
masculinos: caso os masculinos estejam representados em uma coluna os
femininos são representados em uma linha, mas se
os masculinos foram
representados em uma linha os femininos ficam em uma coluna. No diagrama
também visualizamos como resultado do encontro gamético o genótipo e fenótipo
dos descendentes. (fig. 1.11).
27
Figura 1.11: Representação do Cruzamento-teste de uma só característica utilizando o
quadrado de Punnett: Em (a), a planta genitora Cotilédone amarelo é homozigota, mas,
em (b), a genitora Cotilédone amarelo é heterozigota. O genótipo de cada planta alta da
P1 pode ser determinado por meio do exame da prole, quando cada uma é cruzada com
a planta baixa homozigota. recessiva. Adaptada de Klug et. al., 2010.
28
1.9. EXEMPLOS DE HERANÇA MONOGÊNICA OU MONO-HIBRIDISMO EM
OUTROS ORGANISMOS
1.9.1.HERANÇA DO TIPO DE FOLHA EM COLEUS BLUMEI (CÓLEO)
Na planta Coleus blumei, utilizada na ornamentação de jardins, a
característica que determina a forma da borda das folhas : crenada(levemente
ondulada) ou lobadas (profundamente recortada) é determinado por um par de
genes. onde o gene que determina folha lobada é dominante sobre o que determina
folha crenada.(fig. 1.12)
Figura 1.12: Representação esquemática entre plantas de Coleus blumei (foto) de folhas
lobadas e folhas crenadas. Fonte: Amabis, José M.,Martho, Gilberto R., 2006.
29
1.9.2. HERANÇA DO TIPO DE ASA EM DROSOPHILA MELANOGASTER
A mosca Drosophila, também chamada mosca-do-vinagre ou mosca-dabanana também apresenta características determinadas por um único par de
genes, um exemplo é o tipo das asas, onde os fenótipos asa longa (ou selvagem) e
asa vestigial (ou mutante), ao serem cruzados produzem em F 1 toda a prole com
asas longas e em F2 mantém a proporção Mendeliana de 3/4 asas longa para 1/4
asa vestigial. (fig. 1.13)
Figura 1.13: Representação do cruzamento entre Drosophilas selvagens de asas longas
e mutantes de asas vestigiais (na foto, aumento ≈ 17X). Fonte: Amabis, José M.,Martho,
Gilberto R., 2006.
30
1.9.3. HERANÇA DA SENSIBILIDADE AO PTC NA ESPÉCIE HUMANA
Uma herança com padrão de herança monogênica na espécie humana é a
sensibilidade ao PTC (droga denominada feniltiocarbamida, ou feniltiouréia),
pessoas que são sensíveis ao PTC sentem um gosta amargo na boca quando são
colocados em contato com soluções de PTC, e outras pessoas nada sentem quando
em contato com a droga. Sensibilidade ao PTC é dominante em relação à
insensibilidade. (fig. 1.14)
Figura 1.14: Representação esquemática do cruzamento entre uma mulher sensível ao
PTC e um homem insensível ao PTC (Feniltiocarbamida). Adaptado de :Amabis, José
M.,Martho, Gilberto R., 2006.
31
1.10. HEREDOGRAMAS, GENEALOGIAS, ÁRVORES GENEALÓGICAS OU
PEDIGREE
Na espécie humana fica muito difícil determinar se um fenótipo é
hereditário ou não e qual o padrão de herança de uma dada característica, já que
não é possível se fazer cruzamentos experimentais e a prole resultante é muito
pequena, para que se tenha uma boa análise estatística, além disso, a possibilidade
de se estudar o comportamento de um determinado traço por várias gerações em
uma família é pouca provável, já que, às vezes aquele traço só se manifesta na
idade adulta, depois dos 40 anos.
O meio tradicional para o estudo de determinadas características
hereditárias em uma família é a construção de uma árvore familiar ou
heredograma, indicando a presença ou ausência de um traço em questão para cada
membro de cada geração. As genealogias são representações gráficas
convencionadas pelos geneticistas das relações de parentesco entre os indivíduos
de uma família. (fig.1.15)
32
Figura 1.15: Símbolos comumente empregados na representação gráfica de genealogias.
Fonte: Lima, Celso P.,1984
33
1.10.1. ANÁLISE DE GENEALOGIAS
Os heredogramas nem sempre são precisos para diagnosticar o tipo de
herança que estamos trabalhando, já que, o tamanho da amostra é pequeno e a
elaboração muitas vezes depende de informações prestadas pelo probando ou
propósito (isto é, pelo indivíduo afetado que atraiu a atenção dos pesquisadores),
ou por seus parentes que muitas vezes não estão tão interessados em colaborar de
forma mais efetiva, ou ainda, a história da família pode ser falseada porque o
investigador muitas vezes se baseia em dados que depende da memória de quem
os fornece. Para que se possa interpretar com menor chance de erro os dados é
necessário muitas vezes analisar vários genealogias referentes aquele fenótipo que
se está estudando.
Alguns critérios que não são rígidos, mas, que ajudam na identificação de padrões
de herança quando analisamos heredogramas.
Como se reconhece a herança autossômica (característica monogênica onde os
genes encontram-se em cromossomos autossômicos) dominante (fig. 1.16)
1) A característica ocorre igualmente em homens e mulheres;
2) Indivíduos afetados são sempre filhos de casais em que pelo menos um dos
cônjuges é afetado; dessa forma, um casal normal nunca tem filhos afetados
(a não ser por mutação que é raro ou por penetrância incompleta);
3) A característica ocorre em todas as gerações sem pular nenhuma;
Figura 1.16: Heredograma representativo de uma característica autossômica dominante.
Fonte: Klug et. al.,2010
34
Como se reconhece a herança autossômica recessiva (fig.1.17)
1) Os dois sexos são igualmente afetados;
2) Os indivíduos afetados geralmente são filhos de pais normais;
3) Dentre os irmãos do propósito, os indivíduos afetados e normais
distribuem-se na proporção de 3 normais para 1 afetado;
4) Os
indivíduos
afetados
geralmente
resultam
de
cruzamentos
consangüíneos.
Como se reconhece a herança recessiva ligada ao sexo (herança monogênica onde
os genes encontram-se numa porção do cromossomo sexual X que não tem
homologia no Y) (fig.1.18)
1) Mulheres afetadas são muito mais raras do que homens afetados;
2) Homens afetados geralmente têm filhos normais;
3) Os indivíduos afetados são filhos de mulheres normais que, por sua vez, são
filhas de homens afetados; em outras palavras, a anomalia passa de avô
4) para neto, através de suas filhas que são portadoras do gene.
Figura 1.17: Heredograma representativo de uma característica autossômica recessiva.
Fonte: Klug et. al.,2010
35
Figura 1.18: Heredograma representativo de uma característica ligada ao sexo recessiva.
Fonte: Lima, Celso P.,1984.
Como se reconhece a herança dominante ligada ao sexo (fig. 1.19)
1) A característica marcante deste tipo de herança é o fato de que os homens
afetados têm todas as suas filhas afetadas, embora nenhum de seus filhos o
seja;
2) As mulheres heterozigotas transmitem as características à metade de seus
descendentes, sejam meninos ou meninas.
3) As mulheres afetadas homozigotas transmitem as características a todos os
seus descendentes.
4) Este tipo de herança só pode ser reconhecido pela descendência dos
homens afetados; se não existir descendência deles torna- se impossível
reconhecer este tipo de herança, visto que ela se assemelha à herança
autossômica dominante.
36
Figura 1.19: Heredograma representativo de uma característica ligada ao sexo
dominante. Fonte: Lima, Celso P.,1984
1.11. A BASE MOLECULAR DA SEGREGAÇÃO E EXPRESSÃO MONOGÊNICA
1.11.1. COMO SURGEM OS ALELOS!
Quando falamos em alelos até agora não nos preocupamos com a estrutura
e função desses alelos somente como eles se segregam e se comportam em relação
aos outros alelos. Mas, a nível molecular como surgem os diferentes alelos? Esses
alelos são resultantes de mutações de alelos denominados selvagens, que se
caracterizam por aparecer em maior freqüência na população. Os alelos
resultantes de alterações no DNA do alelo selvagem são ditos, alelos mutantes.
Existem vários tipos de mutações, (que serão estudadas em outro capítulo), porém,
as mutações só são visíveis quando elas alteram o gene, de tal forma, que como
conseqüência altere o fenótipo. Um novo fenótipo resulta de uma mudança na
atividade funcional do produto celular (proteína) especificado pelo respectivo
gene.
Esses genes (ou alelos) mutantes podem ser em relação ao alelo selvagem
dominante ou recessivo. Quando a mutação altera o alelo selvagem e ele diminui
ou perde a função essa mutação é denominada mutação de perda de função, se a
perda for completa é formado um alelo denominado alelo nulo e o fenótipo
determinado por esse alelo é geralmente recessivo.
Em outros casos, a mutação altera o alelo selvagem formando alelos
mutantes com um aumento na atividade funcional em relação a atividade funcional
37
do alelo selvagem , aumentando assim, a quantidade do produto gênico, nesse caso,
denominamos a mutação de mutação de ganho de função, e o fenótipo resultante
geralmente é um fenótipo dominante.
Como já havia falado existem várias formas de representação dos alelos, a primeira
mais simples, usada por Mendel, onde se representa o fenótipo dominante com a
letra maiúscula e em itálico, do fenótipo recessivo e o recessivo com letra
minúscula e em itálico. Outra notação, determinada quando Morgan e Bridges
estudavam cor do olho em Drosophila, que pode ser usada para representar o alelo
selvagem e o alelo mutante é a primeira letra ou a combinação de duas ou três
letras do traço mutante em maiúsculo ou minúsculo, quando representa
respectivamente fenótipos mutantes dominantes e recessivos, acrescidos de um
sinal sobrescrito + para representar o alelo selvagem ou simplesmente para
simplificar, o sinal + para o alelo selvagem e o mutante com a letra inicial ou com a
combinação de duas ou três letras. Quando se está representando o genótipo
utiliza-se uma barra separando os alelos de um mesmo locus em cromossomos
homólogos. (fig. 1.20 a e b)
Tabela representando os genótipos e fenótipos para a cor do corpo em Drosophila
que apresenta o alelo mutante recessivo e fenótipo ébano e o alelo selvagem
dominante e+ fenótipo cor cinza.
Fenótipos
Genótipos
e+/e+ ou +/+
Homozigoto cinza (tipo selvagem)
e+/e ou +/e
Heterozigoto cinza (tipo selvagem)
e/e ou e/e
Homozigoto ébano ( tipo mutante
Figura 1.20a: Representação de notação para designar genótipos e alelos selvagens e
mutantes recessivo. Adaptada de Klug et.al., 2010
38
Tabela representando os genótipos e fenótipos para a forma da asa em Drosophila
que apresenta o alelo mutante dominante Wr fenótipo asa rugosa e o alelo selvagem
dominante Wr+ fenótipo asa lisa.
Genótipos
Fenótipos
Wr/Wr
Asa rugosa (tipo mutante)
Wr/Wr+
Asa rugosa (tipo mutante)
Wr+/Wr+
Asa normal (tipo selvagem)
Figura 1.20b: Representação de notação para designar genótipos e alelos selvagens e
mutantes dominantes. Adaptada de Klug et.al., 2010
2. 2ª LEI DE MENDEL OU LEI DA SEGREGAÇÃO INDEPENDENTE
Mendel após trabalhar na observação de uma característica por vez
começou a fazer experimentos com plantas que diferiam em duas características.
Ele cruzou a variedade de plantas que produziam sementes amarelas lisas, e que
por autofecundação só originavam plantas com sementes amarelas lisas, portanto
puras para as características cor e forma da semente, com a variedade de plantas
verdes rugosas também puras para essas características com o objetivo de analisar
como se comportavam as duas características ao mesmo tempo na hora de formar
os gametas e a prole.
Vamos usar as seguintes notações nos cruzamentos para representar os
genótipos que incluem dois pares de alelos que se encontram em cromossomos
homólogos distintos para as características: cor do cotilédone da semente; V
(Amarela) e v (verde) e forma da semente; R (Lisa) e r (rugosa), logo o genótipo da
planta com sementes amarela lisa pode ser assim representado
VV RR e o da
planta verde rugosa vv rr. Ao cruzar essas duas linhagens Mendel obteve em F1
todas as plantas com semente amarelas lisas, indicando que os dois traços, amarelo
e liso são dominantes em relação à verde e rugoso com o seguinte genótipo Vv Rr.
Essas Plantas F1 ele deixou autofecundar e resultou em todas as possibilidades de
combinações nas proporções; 9/16 plantas amarelas lisas, 1/16 plantas verdes
39
rugosas, semelhantes aos fenótipos parentais, 3/16 plantas amarelas rugosas e
3/16 plantas verde lisa. (fig. 1.21 e 1.22)
Figura 1.21: Os cruzamentos de Mendel entre ervilhas que produziram sementes
amarelas e lisas e ervilhas que produziram sementes verdes e rugosas. Fonte: Snustad,
D.Peter; Simmons, Michael J.,2008.
Mendel também usou o cruzamento teste (cruzamento do di-híbrido com o
duplo recessivo) com o intuito de demonstrar que cada gameta do di-híbrido é
formado por um alelo do par V ou v e do par R ou r e que essas combinações
gaméticas apareciam em uma mesma proporção, indicativo de segregação
independente.(fig.1.23)
40
Figura 1.22: Representação simbólica dos resultados de um cruzamento entre uma
variedade de ervilhas com sementes amarelas e lisas e uma variedade com sementes
verdes e rugosas. Fonte: Snustad, D.Peter; Simmons, Michael J.,2008.
41
Figura 1.23: Representação do cruzamento teste de plantas de ervilhas amarela lisa (F 1)
com o duplo recessivo (verde rugosa). Adaptado de Snustad, D.Peter; Simmons, Michael
J.,2008.
Mendel fez outras combinações entre as sete características estudadas por
ele, em todas elas obteve os mesmos resultados e com esses resultados Mendel
chegou às seguintes conclusões:z
1. Cada característica é controlada por um par de alelos que se
segregam na hora de formar os gametas;
2. Cada um dos pares segrega (separa) de forma independente do outro
3. na hora de formar os gametas, logo a alelo V pode estar em um
gameta junto com o alelo R ou com r e o alelo v pode vir em um
gameta tanto junto com o alelo R ou com r, as chances desses
encontros são as mesmas e essas combinações são feitas de forma
aleatória.
42
Após as conclusões, Mendel enuncia o que chamamos 2ª lei de Mendel ou
Lei da segregaç~o independente: “Os alelos de genes diferentes segregam-se, ou
distribuem-se, independentemente uns dos outros.”
Além dos experimentos com uma característica, cruzamentos monohíbridos, ou duas características ao mesmo tempo, cruzamentos di-híbridos,
Mendel fez experimentos com três características ao mesmo tempo, cruzamento
esse chamado tri-híbrido e observou que a segregação independente também é
aplicada nesses casos.
Depois de 1900 quando os trabalhos de Mendel foram redescobertos, as leis
de Mendel foram testadas em várias outras plantas e animais, os resultados
obtidos vieram para validar o trabalho de Mendel quase que de forma geral. A
exceção à lei de segregação independente é quando os dois ou mais pares de alelos
que determinam as características se encontram em um mesmo cromossomo
homólogo.
2.1. A MEIOSE E A 2ª LEI DE MENDEL
Sabemos que em um organismo diplóide cada um dos pares de homólogos é
constituído por cromossomos de origem paterna, proveniente do gameta
masculino e o outro, de origem materna, proveniente do gameta feminino. As
plantas de ervilhas possuem 14 cromossomos, ou seja, 7 pares de homólogos e
Mendel fez experimentos com 7 características ou traços distintos. Os pares de
alelos que determinam cada uma das características mendelianas se encontram
distribuídos nesses cromossomos. Cada uma das características estudada em um
par de homólogos. Os cromossomos homólogos na meiose segregam-se
independentemente levando junto os pares de alelos mendelianos.
Estudos posteriores que analisaram outras características das plantas de
ervilha, não conseguiram obter sempre os mesmos resultados de Mendel a
conclusão para isso é que características que estão em um mesmo par de
cromossomos homólogos segregam juntas na hora de formar os gametas (fig. 1.24)
43
Figura 1.24 Representação esquemática da segregação independente dos cromossomos
homólogos na meiose, responsável pela segregaçáo independente dos genes situados
em diferentes pares de homólogos. Em uma célula duplo-heterozigótica, há duas
possibilidades para a migração dos cromossomos, o que caracteriza a segregação
independente. Fonte: Amabis, José M.,Martho, Gilberto R., 2006.
44
3. APLICAÇÕES DOS PRINCÍPIOS DE MENDEL
Se a base genética de uma característica é conhecida, os princípios de
Mendel podem ser usados para prever o resultado dos cruzamentos. Existem três
procedimentos analíticos que são parte das pesquisas genéticas cotidianas e são
utilizados para se fazer a análise de proporções fenotípicas. Essa análise pode ser
feitas por dois caminhos ou prevendo os genótipos dos genitores a partir das
proporções fenotípicas da prole, ou as proporções fenotípicas da prole tendo-se o
conhecimento dos genótipos dos genitores.
3.1. O QUADRADO DE PUNNETT
O método utilizado até agora “o quadrado de Punnett”, representados na
figura 2.3 e 2.4, é muito útil em situações que envolvem 1 ou 2 pares de genes, pois
dá uma visualização dos gametas formados pelos genitores e a representação de
todos os encontros gaméticos possíveis na formação da prole, resultando nos
genótipos. Podendo-se chegar às proporções fenotípicas quando se sabe a relação
de dominância entre os alelos que compõem os genótipos. Porém se torna muito
trabalhoso usarmos esse método quando passamos a ter 3 ou mais pares de alelos
determinando três ou mais características ao mesmo tempo. Com 3 pares de alelos,
resultantes do cruzamentos de 2 linhagens uma homozigota dominante com uma
homozigota recessiva, teríamos um tri-híbrido em F1. Esse F1 produziria 8 tipos
diferentes de gametas(fig. 1.25) que se for cruzado com outro igual a ele também
formará 8 diferentes tipos de gametas, gerando 8X8 = 64 genótipos, no quadrado
de Punnett.
45
Figura 1.25: Representação esquemática da segregação independente de 3 pares de
alelos originando os oito tipos de gametas. Adaptada de Amabis, José M.,Martho,
Gilberto R., 2006.
3.2. O MÉTODO DA LINHA BIFURCADA
Caracteriza-se por representar os fenótipos ou genótipos resultantes do
cruzamento de cada uma das características estudadas em linhas que se bifurcam,
em cada elo da bifurcação colocam-se os fenótipos ou genótipos de um dos
cruzamentos. (fig. 1.26a e 1.26b)
46
Cruzamento:
Vv Rr Bb
X
Vv Rr
Bb
Segregação de gene Segregação
do Segregação
do Fenótipo combinados de
para cor do cotilédone gene para forma gene para cor da todos os três genes
da semente
casca da semente
da semente
3 Cinzas
27 altas, lisas, cinzas
1 Branca
9 altas, lisas, brancas
3 Cinzas
9 altas,rugosas,cinzas
1 Branca
3 altas,rugosas,brancas
3 Cinzas
9 baixas,lisas,cinzas
1 Branca
3 baixas,lisas,brancas
3 Cinzas
3 baixas,rugosas,cinzas
1 Branca
1 baixa,rugosa,branca
3 Lisas
3 Altas
1 Rugosa
3 Lisas
1 Baixa
1 Rugosa
Figura 1.26a: O método da linha difurcada para prever o resultado de um cruzamento
envolvendo 3 genes que se distribuem independentemente em ervilhas. A proporção
fenotípica é dada pelo produto de cada cruzamento individualmente. Adaptado de
Snustad, D.Peter; Simmons, Michael J.,2008.
47
Cruzamento:
Segregação de gene para cor
do cotilédone da semente
1 VV
2 Vv
1 vv
Vv Rr
X
Vv Rr
Segregação do gene para
forma da semente
1 RR
Proporção dos Genótipos
combinados de 2 pares de
genes.
1 VV RR
2 Rr
2 VV Rr
1 rr
1 VV rr
1 RR
2 Vv RR
2 Rr
4 Vv Rr
1 rr
2 Vv rr
1 RR
1 vv RR
2 Rr
2 vv Rr
1 rr
1 vv rr
Figura 1.26b: O método da linha difurcada para prever o resultado de um cruzamento
envolvendo 2 genes que se distribuem independentemente em ervilhas. A proporção
genotípica é dada pelo produto de cada cruzamento individualmente. Adaptado de
Snustad, D.Peter; Simmons, Michael J.,2008.
Observando as figuras acima vamos perceber que quando montamos a linha
bifurcada para os genótipos fica bem mais difícil manejar. Levando em conta só
duas características di-híbridas, já obtemos 3n onde n é igual ao número de
características heterozigotas e 3 é o número de genótipos distintos em cada
cruzamento mono-híbrido, caso tenhamos 3 pares de genes (cruzamento trihíbrido) teríamos 33= 27 genótipos diferentes.
48
3.3. O MÉTODO MATEMÁTICO
Um método alternativo ao quadrado de Punnett e o da linha bifurcada, e
mais rápido, é baseado no princípio da probabilidade.
Probabilidade é a chance de um determinado evento ocorrer, entre dois ou
mais eventos possíveis. Por exemplo, qual a probabilidade de em um nascimento
obtermos um menino? O número de eventos possíveis (minha amostra) são dois a
criança ou é menino ou é menina. Logo a probabilidade de que seja menino é 1 em
2 ou 1/2.
Eventos aleatórios são eventos que têm a mesma chance de ocorrer
quando comparados com outros eventos possíveis dentro de uma probabilidade.
Eventos independentes são eventos em que a ocorrência de um evento
não afeta a probabilidade do outro evento ocorrer. Exemplo: o nascimento de cada
filho é um evento independente já que nascimento do 1º não afeta ou não interfere
no nascimento do 2º e assim sucessivamente, ou em um cruzamento entre um
casal de heterozigotos, a probabilidade da mulher produzir gametas não interfere
na probabilidade do homem também produzir.
Eventos mutuamente exclusivos são eventos que quando um ocorre o
outro não pode ocorrer ao mesmo tempo. Um bom exemplo dentro da genética
para isso é o nascimento de uma criança, ela não pode ser ao mesmo tempo
menina e menino. Ou ele é menina ou ela é menino. Logo esses eventos são
mutuamente exclusivos.
Em estatística existem 2 regras básicas que são necessárias para a resolução
dos exercícios para cálculo das proporções genotípicas e fenotípicas da prole, são
elas a regra dos produtos (ou regra do “e”) e a regra da soma (ou regra do “ou”).
A regra do produto diz: “A probabilidade de dois eventos independentes
ocorrerem juntos (ao mesmo tempo) é igual ao produto das probabilidades de cada
um deles”. Exemplo: Uma mulher teve 2 crianças, qual a probabilidade que a
primeira seja menina e o segunda seja menino? Em cada nascimento a
probabilidade de ser menino ou de ser menina é a mesma, 1/2, e o nascimento de
cada filho é um evento independente já que nascimento do 1º não afeta ou não
49
interfere no nascimento do 2º. Logo a probabilidade da mulher ter o 1º menina e o
2º menino é igual ao produto das probabilidades individuais (1/2X1/2=1/4)
A regra da soma diz: “A probabilidade de dois eventos mutuamente exclusivos
ocorrerem é igual a soma das probabilidades de cada um ocorrer”.Exemplo; Qual a
probabilidade de em um nascimento nascer uma menina ou um menino? A
probabilidade é a soma das probabilidades individuais (P(menino=1/2))+
(P(menina=1/2)) = (1/2 + 1/2)= 1.
3.3.1.
USANDO
AS
REGRAS
DE
PROBABILIDADE
PARA
REALIZAR
CRUZAMENTOS.
1. Em um cruzamento monogênico, qual a probabilidade de obtermos
indivíduos heterozigotos do cruzamento de um casal de heterozigotos
com genótipos Aa?
Cruzamento:
♀
♂
A
1/2
a
1/2
Aa
X
Aa
A
a
1/2
1/2
AA
aA
1/2x1/2=1/4 1/2x1/2=1/4
Aa
Proporção
genotípica
Proporção
fenótípica
1/4 AA
1/4+2/4 = 3/4 A_
Fenótipo
dominante
1/4 + 1/4 Aa= 2/4
1/4 = aa
aa
1/2x1/2=1/4 1/2x1/2=1/4
1/4 = aa
Fenótipo
recessivo
50
2. Do cruzamento de duas plantas com genótipos Aa BB Cc Dd Ee X Aa Bb
Cc dd Ee qual a probabilidade de obtermos na prole um descendente
com o seguinte genótipo:
aa Bb Cc Dd ee
Resposta: Como cada característica segrega independente (são
eventos independentes); já que estamos considerando que se encontram
em cromossomos homólogos diferentes;
poderemos achar a
probabilidade individual em cada cruzamento Aa X Aa. BB X Bb, Cc X Cc,
Dd X dd e
Ee X Ee e depois multiplicar cada uma das probabilidades.
Aa X Aa = 1/4 aa; BB X Bb = 1/2 Bb; Cc X Cc = 1/2 Cc; Dd X dd = 1/2 Dd;
Ee X Ee = 1/4 ee.
P( aa Bb Cc Dd ee)= P(aa)XP(Bb)XP(Cc)XP(Dd)XP(ee)=
1/4X1/2X1/2X1/2X1/4= 1/128
3.3.2 QUANTOS GENÓTIPOS DISTINTOS UM CRUZAMENTO PRODUZ?
As regras de probabilidades podem ser facilmente utilizadas para prever
quantos genótipos ou fenótipos diferentes podem surgir na prole de linhagens
parentais complexas com quatro cinco ou mais pares de genes. Exemplo: No
cruzamento de tetra-híbrido quantos genótipos e quantos fenótipos diferentes
podem ter? Cruzamento tetra-híbrido: Aa Bb Cc Dd X Aa Bb Cc Dd. Cada
cruzamento individual gera 3 genótipos diferentes AA, Aa, aa e dois fenótipos
diferentes o dominante e o recessivo.podemos utilizar a fórmula 3n onde n é o
número de características individuais, ou seja o número de genótipos diferentes é:
34 = 81; já o número de fenótipos é 2n= 24 = 16.
Em um cruzamento teste Aa Bb Cc Dd X aa bb cc dd; cada cruzamento
individual produz 2 tipos de genótipos o Aa e o aa e também dois tipos de
fenótipos o dominante e o recessivo, nesse caso a mesma fórmula pode ser usada
tanto para calcular o genótipo quanto o fenótipo 2 n = 24 = 16
51