USO DE FONTES DE CARBONO POR BACTÉRIAS ISOLADAS DE ARROZ
(Oryza sativa L.)
2
Lucas L. da Silva*¹ (PG), Larissa B. da Silva¹(PG), Enderson P. B. Ferreira (PQ), Karina F. D. N.
Santos¹(PQ), Plínio L. F. Naves¹(PQ), Claudia C. G. Martin-Didonet¹(PQ).
¹Br 153, Nº3105 Fazenda Barreiro do Meio, Câmpus de Ciências Exatas e Tecnológicas – CCET,
Anápolis, GO – [email protected]
²Empresa Brasileira de Pesquisa Agropecuária Arroz e Feijão, Santo Antônio de Goiás, GO.
Resumo: O objetivo desse estudo foi caracterizar o crescimento bacteriano quanto à utilização de
diferentes fontes de carbono em 20 bactérias associadas à planta de arroz (Oryza sativa L.). A
capacidade de metabolizar diferentes fontes de carbono foi determinada por espectrofotometria a 600
nm, sendo os resultados convertidos em dados binários e utilizados para construção de um
dendrograma. Mediante as características dos isolados foi observado que 35% dos isolados
metabolizaram entre oito a nove das fontes testadas e 87% não foi capaz de crescer em meio
enriquecido com ácido maléico. Pelo dendrograma foram identificados nove grupos com índice de
similaridade em torno de 80%, sendo quatro isolados 100% similar às estirpes de referência BR322 e
BR520. Assim, a análise permitiu a caracterização de isolados de raiz de arroz, os quais
apresentaram alta diversidade metabólica frente às fontes de carbono testadas.
Palavras-chave: Diversidade metabólica. Caracterização fenotípica. Micro-organismos.
Introdução
Os micro-organismos podem ser encontrados em diversos habitats como o
solo. Este é de grande heterogeneidade física, química e biológica e representa um
dos maiores reservatórios de diversidade microbiana, sendo fonte para a exploração
biotecnológica. Dentre os micro-organismos presentes no solo, várias espécies de
bactérias podem viver livremente ou estarem associadas às plantas, estabelecendo
uma importante relação planta-solo-micro-organismos (CARDOSO e NOGUEIRA,
2007; ARAÚJO, 2011; MARON et al., 2011; KUJUR et al., 2012).
As bactérias endofíticas podem colonizar o interior dos tecidos de alguns
vegetais sem causar danos por sua presença, e têm sido mencionadas em vários
estudos como rizobactérias promotoras do crescimento de plantas ou Plant-Growth
Promoting Rhizobacteria (PGPR). Alguns grupos dessas bactérias são capazes
de fixar biologicamente o nitrogênio (FBN) atmosférico (N2) em íon amônio (NH4+),
sendo denominadas de diazotróficas. O nitrogênio é um elemento fundamental para
o desenvolvimento das plantas, sendo que as bactérias diazotróficas endofíticas
podem fornecer total ou parcialmente as necessidades deste composto (VARGAS e
HUNGRIA, 1997; ESPOSITO-POLESI, 2011).
Alguns trabalhos têm demonstrado a diversidade microbiana através de
caracterizações fisiológicas tais como a capacidade de metabolizar diferentes fontes
de carbono (ALVES, 2005; RODRIGUES et al., 2006; BARBOSA et al., 2006;
CASTELLANE e LEMOS 2007; LEONEL et al., 2014) . Nesse contexto, o objetivo
desse estudo foi caracterizar o crescimento bacteriano quanto à utilização de
diferentes fontes de carbono a partir de 20 bactérias associativas de arroz (Oryza
sativa L.).
Material e Métodos
Material Biológico: As bactérias utilizadas neste estudo foram isoladas de raízes de
plantas de arroz e mantidas no Banco de Micro-organismos do Laboratório de
Bioquímica-UEG. Os isolados foram testados quanto á sua capacidade em
metabolizar diferentes fontes de carbono (C).
Meio de cultura e teste de fontes de carbono: As bactérias foram cultivadas em
caldo semi-seletivo extrato de levedura-manitol (YM) para bactérias da família
Rhizobiaceae, por 24h, sob agitação (140 rpm) a 25 ºC e utilizados 50µL nos testes
de fontes de C (ácido maléico, ácido málico, ácido succínico, frutose, glicose,
inositol, manitol, manose, sorbitol e trealose) como pré-inóculo (VARGAS e
HUNGRIA, 1997). Para o teste foi utilizado o meio mínimo líquido (HUNGRIA et al.,
1994) e as fontes de C foram adicionadas por filtração na concentração de 10 mM e
distribuídos (1 mL) em micro placas de 96 poços (deep well plate) para crescimento.
Estas foram incubadas sob agitação (140 rpm) a 25ºc por 72h. O crescimento celular
foi avaliado por espectrofotometria (Epoch®) a 600 nm.
Análise dos dados: Através das médias de leitura de cada isolado, os resultados
foram convertidos em dados binários e utilizados para construção de uma matriz,
adotando os critérios de faixas de leituras. A partir deste processo foi derivada uma
matriz de similaridade utilizando o coeficiente de Jaccard (J) para a construção do
dendrograma, por intermédio do algoritmo UPGMA (Unweighted Pair Group with
Arithmetic Means), através do software NTSYS-pc® 2.10 (RODRIGUES et al.,
2006).
Resultados e Discussão
Mediante as características dos isolados e das estirpes de referência quanto à
capacidade em metabolizar diferentes fontes de C foi observado que 35% dos
isolados metabolizaram entre oito a nove das fontes testadas, 87% não foi capaz de
crescer em meio enriquecido de ácido maléico, e 9% apresentou crescimento em
apenas uma fonte. Dentre as fontes mais utilizadas estão a glicose, inositol, sorbitol
e trealose, e as mais discriminadas o ácido maléico, ácido málico e frutose.
Resultados semelhantes foram observados por Sampaio (2015) em estudo realizado
com bactérias do solo isoladas no estado de Goiás.
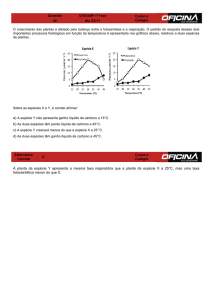
Pela análise dos resultados foi elaborado um dendrograma sendo definidos
nove grupos com índice de similaridade em torno de 80% (Figura 1). Entre os
agrupamentos, G1 e G2 foram compostos por dois isolados cada: R18, R139 e R20,
R64 respectivamente. Ambos os grupos mostraram crescimento insatisfatório e/ou
ausente em meio contendo frutose como fonte de carbono.
O maior agrupamento obtido (G4) apresentou 100% de similaridade com as
estirpes de referência BR322 e BR520 (Rhizobium tropici), com exceção do isolado
R21 (Figura 1). As bactérias inclusas nesse grupo demonstraram ser capaz de
assimilar todas as fontes de carbono fornecidas, indicando semelhanças
metabólicas entre as mesmas. Já os isolados R136a, R148, R161 (grupo G5) e R23,
R59, R141, R50 R68 (grupo G6) mostraram 70% de similaridade com as estirpes de
referência.
Em relação aos demais isolados, os grupos G7 (R62), G8 (R130) e G9
(R145), foram os que apresentaram resultados mais divergentes não agrupando
com nenhum outro grupo. Foi observado que entre as fontes testadas os isolados
R62 e R145 apresentaram crescimento favorável em apenas uma fonte de C
(inositol e trealose, respectivamente).
Alguns estudos têm elucidado o uso de diferentes carboidratos (glicose,
sacarose, frutose, arabinose) e sais de ácidos orgânicos como fontes de carbono por
bactérias do gênero Rhizobium, sendo esta habilidade relacionada à disputa por
nichos na rizosfera (GRAHAM et al., 1991; CAVAGLIERI et al., 2004; CASTELLANE
e LEMOS 2007). Os resultados indicam que os isolados de arroz podem estar
taxonomicamente relacionados à família Rhizobiaceae pela semelhança metabólica
observada, porém outras características são necessárias para determinação
taxonômica das amostras. Muitos autores expõem a importância da caracterização
fenotípica para seleção de micro-organismos com potencial aplicação biotecnológica
e
direcionamento
taxonômico,
sendo
comumente
empregados
parâmetros
ecológicos, sorológicos, resistência a antibióticos e tolerância a diversos fatores, tais
como: salinidade, variações do pH e temperatura, uso de fontes de carbono dentre
outros (PEREIRA, 2002).
Coefficient Jaccard
Figura 1: Dendrograma de similaridade gerado pelo software NTSYS-pc® 2.10 com base no
uso de fontes de carbono (ácido maléico, ácido málico, ácido succínico, frutose, glicose,
inositol, manitol, manose, sorbitol e trealose), definido pelo algoritmo UPGMA utilizando o
coeficiente de Jaccard.
Conclusão
A análise permitiu a caracterização de isolados de raiz de arroz, os quais
apresentaram alta diversidade metabólica frente às fontes de carbono testadas. O
maior grupo formado (G4), apresentou 100% de similaridade com as estirpes de
referência (Rhizobium tropici), apresentando também a maior capacidade de
assimilação de fontes alternativas de C.
Agradecimentos
Ao fomento fornecido pela Fundação de Amparo à Pesquisa do Estado de Goiás (FAPEG),
pela bolsa em Pesquisa de Pós-graduação nível mestrado Stricto Sensu do Programa de Mestrado
em Ciências Aplicadas a Produtos para Saúde. Ao Instituto Nacional de Ciência e Tecnologia (INCTCNPq) em Fixação Biológica de Nitrogênio pelo financiamento do projeto.
Referências
ALVES, J. B. Seleção de rizóbios para trevo branco. 2005. 68 f. Dissertação
(Mestrado em Agronomia). Faculdade de Agronomia, Universidade Federal do Rio
Grande do Sul, Porto Alegre, 2005.
ARAÚJO, J. F. Diversidade bacteriana do solo em diferentes fitofisionomias do
Bioma Cerrado e perspectivas biotecnológicas. 2011. 153 f. Tese (Doutorado em
Ciências Genômicas e Biotecnologia), Universidade Católica de Brasília, Brasília,
2011.
BARBOSA, E. A.; PERIN, L.; REIS, V. M. Uso de diferentes fontes de carbono por
estirpes de Gluconacetobacter diazotrophicus isoladas de cana-de-açúcar. Pesquisa
Agropecuária Brasileira, v. 41, n. 5, p. 827-833, 2006.
CARDOSO, E. J. B. N.; NOGUEIRA, M. A. A rizosfera e seus efeitos na comunidade
microbiana e na nutrição de plantas. In: Silveira, A. P. D.; Freitas, S. S. (org.).
Microbiota do solo e qualidade ambiental. Campinas: Instituto Agronômico, p.7896, 2007.
CASTELLANE, T. C. L.; LEMOS, E. G. M. Composição de exopolissacarídeos
produzidos por estirpes de rizóbios cultivados em diferentes fontes de
carbono. Pesquisa Agropecuária Brasileira, p. 1503-1506, 2007.
CAVAGLIERI, L.; PASSONE, A.; ETCHEVERRY, M. Screening procedures for
selecting rhizobacteria with biocontrol effects upon Fusarium verticillioides growth
and fumonisin B1 production. Research in Microbiology, v.155, p.747-754, 2004.
GRAHAM, P.H.; SADOWSKY, M. J.; KEYSER, H. H.; BARNET, Y. M.; BRADLEY, R.
S.; COOPER, J. E. ; DE LEY, D. J.; JARVIS, B. D. W.; ROSLYCKY, E. B.;
STRIJDOM, B. W.; YOUNG, J. P. W. Proposed minimal standards for the description
of new genera and species of root - and stem - nodulating bacteria. International.
Journal of Systematic Bacteriology, vol. 41, n. 4, p. 582-587, 1991.
HUNGRIA, M. Metabolismo do carbono e do nitrogênio nos nódulos. In: HUNGRIA,
M. & ARAÚJO, R., eds. Manual de métodos empregados em estudos de
microbiologia agrícola. Embrapa, Brasília, p. 890, 1994.
KUJUR, M.; GARTIA, S.K.; PATEL, A.K. Quantifying the contribution of different soil
properties on enzyme activities in dry tropical ecosystems. ARPN Journal of
Agricultural and Biological Science, v.7, p. 763-772, 2012.
LEONEL, T. F.; MORETTO, C.; MENECHINI, A. K.; LEMOS, E. V. G. M. Sacarose
como fonte de carbono produção de exopolissacarídeos por mesorhizobium
spp. Ciência & Tecnologia, v. 6, n. 1, 2014.
MARON, P. A.; MOUGEL, C.; RANJARD, L. Soil microbial diversity: Methodological
strategy, spatial overview and functional interest. Comptes Rendus Biologies, v.
334, p. 403-411, 2011.
RODRIGUES, L.; BALDANI, V. L. D.; REIS, V. M.; BALDANI, J. I. Diversidade de
bactérias diazotróficas endofíticas dos gêneros Herbaspirillum e Burkholderia na
cultura do arroz inundado. Pesq. agropec. bras., Brasília, v. 41, n. 2, p. 275-284,
2006.
SAMPAIO, F. B. Isolados de rizóbios capturados por genótipos silvestres de
feijoeiro: obtenção, morfologia e uso de fontes de carbono. 2013. 89 f.
Dissertação (Mestrado). Universidade Federal de Goiás, Goiânia, 2013.
VARGAS, M. A. T.; HUNGRIA, M. Fixação biológica do nitrogênio em soja. In:
VARGAS, M. A. T.; HUNGRIA, M. (ED.). Biologia dos solos dos cerrados.
Planaltina, DF: EMPRAPA-CPAC, p. 295-360, 1997.
ESPOSITO-POLESI, N. P. Microrganismos endofíticos e a cultura de tecidos
vegetais: quebrando paradigmas. R. bras. Bioci., Porto Alegre, v. 9, n. 4, p. 533-541,
out./dez. 2011.
PEREIRA, K. C. Caracterização morfológica, bioquímica e molecular de rizóbios
recomendados para inoculação de leguminosas arbóreas. 2002. 74f.
Dissertação (Mestrado para obtenção do titulo de Mestre em Microbiologia).
Universidade Estadual Paulista, São Paulo. 2002.