
FeSBE
Federação das Sociedades de Biologia Experimental
SBNeC
Sociedade Brasileira de Neurociências e Comportamento
Comportamento de peixes: Vantagens e utilidades nas
neurociências
Amauri Gouveia jr
Colaboram:
Caio Maximino
Thiago Marques de Brito
2006
2
Reprodução permitida desde que citada fonte
Como citar este trabalho em referências:
Gouveia Jr, A., Maximino, C. & Brito, T.M. (2006). Comportamento de peixes:
Vantagens e utilidades nas neurociências. Faculdade de Ciências/UNESP.
Bauru: SP.
DIVISÃO TÉCNICA DE BIBLIOTECA E DOCUMENTAÇÃO
UNESP – BAURU
G739c
Gouveia Jr, Amauri.
Comportamento de peixes : vantagens e utilidades nas neurociências /
Amauri Gouveia Junior, Caio Maximino, Thiago Marques de Brito. –Bauru : [s.n.], 2006.
80 f.
1. Peixes. 2. Comportamento. 3. Modelos animais. 4. Ansiedade. I.
Maximino, Caio. II. Britto, Thiago Marques de. III. Título.
CDD 591.5
Ficha catalográfica elaborada por Maristela Brichi Cintra – CRB 5046
3
“O peixe é bastante diferente
Ninguém pode entender como é seu gênio
Reservas porções de oxigênio
Mutações para o meio ambiente
Tem mais cartilagem resistente
Habitando na orla ou profundeza
Devora outros peixes pra despesa
E tem época do acasalamento
revestido de escamas esse elemento
Com a força da santa natureza”
(Ivanildo Vilanova e Xangai - Natureza (trecho))
4
sumário
Prefácio ............................................................................... 04
Cap 1 - Vantagens do uso de peixes ............................... 05
Cap 2 – O que é comportamento?.................................... 13
Cap 3 – Modelos comportamentias em peixes ............... 19
Cap 4 – Modelos de ansiedade piscina .......................... 45
Referências ........................................................................ 57
Anexo I – modelo de laboratório portátil para
comportamento de peixes ................................................ 71
5
Prefácio
Os estudos comportamentais com peixes possuem uma longa tradição que
remonta ao advento da psicologia científica. Thorndike (1911) utilizou a espécie
Fundulus sp em estudos de labirinto. Lorenz (1983) executou diversos estudos
etológicos com peixes cliclídeos. Peixes como o Danio rerio (Paulistinha,
Zebrafish) são utilizados como modelos de desenvolvimento e bioindicadores.
A fisiologia e a ecologia de peixes são assuntos que mobilizam
centenas de laboratórios no mundo e os estudos relativos a aqüicultura são
fonte de crescente interesse e financiamento. No entanto, o estudo do
comportamento de peixes não desperta tanto interesse; Volpato (2003 –
comunicação pessoal) levantou apenas 19 grupos brasileiros que trabalhavam
com comportamento de peixes. Destes dados, podemos concluir que o uso de
peixes como modelos comportamentais animais não é uma coisa comum.
O objetivo deste texto é divulgar algumas das possibilidades de uso e
facilidades a partir de uma revisão da literatura que, ainda que não seja exaustiva,
possa servir de incentivo para que mais grupos se animem a este tipo de pesquisa
e permitam o aumento da comunidade e a densidade de pesquisadores na área.
Amauri Gouveia Jr
6
Cap. 1
Vantagens do uso de peixes
Peixes como animais experimentais
Peixes, de forma geral, podem ser definidos como um conjunto de espécies
que vivem em ambiente aquático, possuem escamas, utilizam-se de movimentos
da base da calda para locomoverem-se e tem o sistema olfativo diferenciado da
respiração (Pough, Janis & Heiser, 2003); as semelhanças entre os diversos taxa
param aí.
Algumas espécies apresentam sensores químicos ou físicos especializados
para determinados estímulos, como campos elétricos; outras podem gerar campos
elétricos com funções diversas de eletrorrecepção e eletrocomunicação; algumas
apresentam visão de cores, outras têm olhos residuais ou mesmo ausentes.
Embora grande parte deles absorva oxigênio da água pelas guelras ou pele, tal
função não é regra. Os ambientes em que são encontrados variam desde fossas
abissais até poças de água, sob uma diversidade de condições de salinidade,
dureza da água, pH, turbidez, ou quantidade de matéria em suspensão.
Sua distribuição comportamental em termos de alimentação inclui desde
filtradores de resíduos até carnívoros, passando por diversos tipos de dieta.
Existem peixes gregários, solitários, que estabelecem hierarquias, que formam
casais e que constroem ninhos. Seu tamanho pode variar desde milímetros até 7
7
metros; sua reprodução pode ser por partenogênese, ovípara ou ovovivípara;
pode apresentar cuidado com o filhote ou não. Tal diversidade, antes de constituirse em um obstáculo, é uma vantagem para seu uso como animal de laboratório e
seu estudo.
Talvez a primeira coisa a se considerar é que pensar em peixes como uma
classe homogênea é uma abstração. Um tubarão tem tanto a ver com uma enguia
como um ornitorrinco com um elefante. Desta forma, falar de peixes de forma
genérica é sempre um risco.
O primeiro conselho para quem se aventurar no estudo comportamental de
peixes é: conheça o “seu” animal. A ecologia, formas de manutenção, hábitos,
alimentação e fisiologia devem ser exaustivamente estudadas na literatura embora para a grande maioria dos peixes isto não possa ser feito, em especial
para os peixes neotropicais.
Há cerca de dois anos atrás, um conhecido pesquisador de peixes
neotropicais de Manaus declarou que a fisiologia de peixes era por vezes divisível
em “trutologia” e o “resto”, dado o grande número de estudos com estes
salmonídeos e a tendência dos pesquisadores de generalizarem seus dados para
outras espécies; no entanto, a biologia de peixes tropicais é uma área de grande
variabilidade entre espécies, fruto de um enorme processo de especialização e
especiação em ambientes dos mais variados que levaram ao desenvolvimento de
adaptações bioquímicas e fisiológicas únicas, que tornam este estudo ainda mais
fascinante (Val e Almeida-Val, 1999).
Os peixes são divididos taxonomicamente, de forma inicial, em agnatos e
gnatotomatos, conforme a ausência ou presença de mandíbulas, respectivamente,
8
sendo a primeira classificação corresponde a lampreias e feiticeiras e a segunda a
todos os demais peixes.
A segunda categoria (os gnatotomatos) é tradicionalmente dividida em
elastomobranchios e hosteochitideos, sendo os primeiros correspondentes aos
peixes cartilaginosos (tubarões, raias e quimeras) e os segundos a todos os
demais peixes ósseos. Estes últimos são divididos em sarcopterigios e teleósteos.
A figura 1, abaixo, apresenta a divisão taxonômica dos peixes. Os peixes
mais utilizados em pesquisa comportamental são teleósteos; isto se deve à sua
maior dispersão geográfica e conhecimento de sua fisiologia. Dentre estes, os
peixes utilizados em aquarismo são mais utilizados, provavelmente devido a seu
manejo já estabelecido, facilidades de reprodução (em parte deles, especialmente
aqueles de domesticação mais antiga, como a carpa e o goldfish) e de
manutenção. Pelos mesmos motivos, observamos predomínio dos peixes de
água-doce.
Em geral, os peixes utilizados em pesquisa laboratorial são de pequeno
porte, de forma que se pode montar um laboratório em espaços reduzidos (eu
mesmo tive o meu primeiro laboratório de peixes em minha sala no departamento
onde dou aulas, sobre uma mesa de não mais de 1,60 x 0,80 m).
9
figura 1 – distribuição taxonômica dos peixes. (Adaptado de Emde, 2000).
10
Além disto, os animais podem ser mantidos em grupos relativamente
grandes (de até 15 a 25 por aquário) de forma tranqüila. Sua manutenção exige
alguns cuidados básicos, o que, por sua vez, exige a compra de alguns
equipamentos (um termostato, lâmpada, um filtro externo ou aerador e reagentes
para a qualidade de água - sempre lembrando que peixes gostam de água com
uma biota adequada, não tão limpa) - mas estes são de baixo custo quanto
comparados com os exigidos por animais experimentais mais tradicionais, como
roedores.
Peixes também são animais que consomem, proporcionalmente, menos
comida que mamíferos, o que reduz os gastos com alimentação. Estes
equipamentos e formas de manejo mais simples podem ser encontrados em bons
livros de aquarismo e comprados em petshops.
Um modelo de laboratório portátil para o estudo de comportamento de
peixes encontra-se no anexo 1.
Tabela 1 – equipamentos básicos para manutenção de peixes.
•
•
•
•
•
•
•
Aquários (podem ser substituídos por potes plásticos, caixas d’água e similares)
Areador (para quebrar a tensão superficial da água e permitir troca gasosa)
Filtro (externo ou biológico)
Termostato (adequar a potência ao tamanho do aquário)
Luzes
Temporizador (“timer”)
Testes e corretores para:
o pH (pHmetro, se houver)
o Nitritos (para aquários marinhos)
o Amônia
o Dureza da água
Os aquários podem ser de plástico, vidro ou acrílico. Sua opacidade e
forma podem variar grandemente segundo os objetivos do ambiente: reprodução,
11
manutenção ou criação, bem como pela espécie de peixe que se cria. Em geral,
tanques e aquários maiores permite maior estabilidade de ambiente e
conseqüentemente, menor perda de sujeitos e cuidados com a manutenção.
A Tabela 1 indica os equipamentos básicos para manutenção dos animais.
Se esta for à linha principal do laboratório, e irá ser mantida por muito tempo,
sugere-se que os equipamentos sejam adaptados para grande porte, com a
substituição, por exemplo, de aeradores por turbinas de ar ligadas a tubos.
As espécies a serem utilizadas em laboratórios piscinos podem variar
grandemente,
segundo
os
objetivos
das
pesquisas,
possibilidade de coleta, compra ou reprodução destas.
espaço
disponível,
Em uma revisão não
exaustiva da literatura que usaremos aqui, as principais espécies estudadas para
comportamento são apresentadas na tabela 2.
Cabe destacar que o Goldfish (Carassius auratus) corresponde a quase
50% de todos os trabalhos, seguido pelo Paulistinha (Zebrafish, Danio rerio - 25%)
e pela tilápia (Oreochromis niloticus -15 %).
Tabela 2 – Espécies estudadas em aspectos comportamentais na revisão
apresentada
• Betta splendens
• Carassius auratus
• Cichlassoma meeki
• Fundulus heteroclitus
• Hexagrammus decagrammus
• Macropodus opercularis
• Oncorhicus mykiss
(anteriormente Salmo
gairdneri)
• Pachyurus squalmipinnis
• Rivulus marmuratus
• Thymallus thymallus
•
Brachyhypopomus pinnicaudatus
• Danio rerio
• Gymnotus carapo
• Ictalurus nebulosus
• Notemigonus crysoleuca
• Oreochromis niloticus
• Poecilia reticulata
• Sciaenops ocellatus
• Xyphophorus variatus
• Micropterus salmoides
12
Chama a atenção à pequena participação de peixes amazônicos,
representados pela Tuvira ou carapó (Gymnotus carapo) e pelo Lebiste ou guppy
(Poecilia reticulata). Esta concentração em poucas espécies tem motivos
históricos (o goldfish foi um dos primeiros peixes domesticados), de disseminação
(o guppy é peixe introduzido em vários países), de uso de dados “secundários” (o
zebrafish é um dos modelos de embriologia mais utilizados no mundo; a Tilapia é
o principal peixe de aqüicultura tropical) ou por propriedades dos peixes (a Tuvira
é um peixe fracamente elétrico).
Claro que o uso de peixes não se limita a estudos comportamentais. Em
uma revisão (Bollis et al., 2001), encontram-se diversos usos de peixes como
animais
experimentais:
fisiologia
cardíaca
e
respiratória;
cultura
celular;
ecotoxicologia; endocrinologia; carcinogênese; gerontologia; neurofarmacologia; e
biologia molecular. Bollis et al. (2001) indicaram a possibilidade do uso de
modelos comportamentais animais em 6 destas categorias. Apesar disto, os
objetivos deste texto se concentrarão em modelos comportamentais.
Conforme foi falado, a revisão feita não é exaustiva e outros artigos que
apresentam o mesmo modelo não foram citados. Desta forma, estudiosos de
agressão, por exemplo, vão nos acusar de poucos artigos deste modelo, mas o
objetivo é simplesmente dar uma geral das formas de trabalhar em
comportamento mais comuns, com ênfase em modelos de ansiedade.
13
Cap 2
O que é comportamento?
Em geral o uso do termo “comportamento” tem significados diferentes para
diferentes autores; grosso modo, pode ser agrupado em dois significados básicos:
é comportamento todo ato motor de um animal; é comportamento todas as
interações de um animal com seu ambiente, como organismo, e não somente os
atos motores.
Esta segunda definição apresenta duas acepções básicas: por um lado, o
comportamento é uma espécie de “fisiologia de segunda ordem”, que se refere
não à fisiologia do sistema ou de parte dele, mas do conjunto de sistemas, e assim
inclui tudo aquilo que normalmente é caracterizado como “psicológico” - como
pensamento, linguagem, memória, percepção, etc 1 ; por outro, as unidades
definidas como alvo de estudo do comportamento podem ser maiores ou menores,
segundo quem as define.
1
Isso não implica, entretanto, que as “leis psicológicas” são redutíveis às “leis do comportamento”.
14
De forma geral, quando um biólogo fala de comportamento, fala de grandes
unidades, como o comportamento alimentar ou reprodutivo; um psicólogo, por
outro lado, fala de unidades menores, como apertar uma barra ou caminhar.
Estas definições são, sem sombra de dúvida, bastante controversas, e
geraram uma grande quantidade de estudos que fogem do escopo deste breve
trabalho. 2 O quadro 1 apresenta níveis de organização em que o comportamento
pode ser estudado. Uma breve olhada sobre este indica que o comportamento
pode ser tanto algo coletivo, como a ação de parte do corpo. Tal variabilidade de
usos de termos exige que se defina claramente o que está sendo chamado de
“comportamento” para o julgamento das estratégias utilizadas.
A pesquisa comportamental pode ser de dois tipos: centrada em espécies,
ou centrada em comportamentos.
No primeiro tipo de pesquisa, em geral
identificada com a zoologia e etologia clássica, bem como com a ecologia
comportamental, o objetivo é conhecer o particular de um comportamento de uma
espécie, e, comumente, compará-lo com o de espécies próximas ou de ecologia
similar.
No segundo tipo, o objetivo é a dedução de uma lei geral de
comportamento, válida para um grande número de espécies - em geral, tal tipo de
pesquisa é identificada com o behaviorismo e as ciências cognitivas, bem como
com a psicologia cognitiva.
Normalmente, o primeiro grupo de pesquisa visa a busca de causa
evolutivas para o comportamento, relacionando as respostas comportamentais a
2
Estudos de campo em geral são referentes aos seguintes aspectos da vida dos animais: alimentação; cuidado
com a prole; mating; comportamento agonistico e hierarquia; organização de grupo e migração.
15
particularidades da seleção natural ou fatores motivacionais intrínsecos; o
segundo grupo de pesquisas busca a dedução de leis gerais do comportamento,
em geral ligadas a aprendizagem e memória, sem muita ênfase sobre as
características espécies especificas destes comportamentos (Lehner, 1996).
O conjunto de elementos que apresentamos tanto é valido para o estudo
das bases fisiológicas do comportamento, como para sua organização mais
macro; tanto para uma situação natural, como para o laboratório; tanto para o
estudo de espécies e grupos, como para o estudo de indivíduos, gerando um
modelo tridimensional do estudo do comportamento.
Quadro 1 – Níveis de organização do comportamento que podem ser alvo de
pesquisas (inspirado em Lehner, 1996).
Nível de organização
Exemplo
Espécie
Fatores sazonais que determinam a migração de peixes oceânicos
População
Comportamento migratório de peixes
Grupos familiares
Cuidado parental no Betta splendens
Díades
Display de agressão no Peixe-do-paraiso (Macropodus opercularis)
Indivíduos
Busca alimentar
Tipo de comportamento Alimentação
Ato comportamental
Mordida
Partes do corpo
Mandibula
Músculos
Cardíaco
Neurônios/nervos
Vago
Bioquimica
Variação de 5-Ht
Para nossos propósitos, vamos definir como comportamento tudo o que o
animal faz, e assumiremos que a unidade de análise pode ser variada e será
explicitada sempre que possível. Por outro lado, não assumimos que fenômenos
16
como memória, percepção ou outros podem ser descritos sem estes termos
específicos ou sejam redutíveis a uma única natureza de fenômeno, mas
assumiremos que, dentro do repertório de coisas que um animal pode fazer,
existem diversas categorias funcionais que podem ser estudadas de formas
diversas ou complementares (cf. Donahoe & Palmer, 1994).
Em geral, a análise de um comportamento, como definido acima, implica
em manipulação de variáveis biológicas ou ambientais e a mensuração de um
padrão de ação definido. Por exemplo, sabemos que ratos expostos a uma
situação composta por uma arena circular tendem a distribuir sua exploração de
forma desigual entre o centro deste aparato e a periferia, explorando mais esta
última.
Sabemos também que o comportamento de explorar (isto é, andar pelo
aparato) diminui ao longo do período de exposição. Neste caso, variações
ambientais de luminosidade, tamanho do aparato, horário do dia em que é feito o
teste, cor do aparato, e objetos introduzidos na arena alteram este padrão de
ambulação. Sabemos também que drogas, lesões, privação de água ou alimento,
e variações de cepas, entre outras variáveis, alteram o comportamento (cf. Prut &
Belzung, 2003).
O cruzamento entre estas variáveis é objeto de estudo nosso. Pode-se
também analisar como a ambulação, ou outros comportamentos emitidos pelo
sujeito experimental, se dá. É claro que o comportamento não ocorre ou não pode
ser somente estudado em situação laboratorial, mas para os nossos propósitos
somente nos deteremos neste tipo de estudo.
17
Resumidamente, o estudo experimental do comportamento pode ter três
objetivos diferentes: o comportamento pode ser uma medida indireta de um efeito
(como nos modelos que aqui falamos, por vezes chamados de ensaios
biocomportamentais ou bioensaios comportamentais); pode ser um estudo de
seus elementos determinantes (tanto ambientais como biológicos) em termos
paramétricos (em geral , o objeto das ciências do comportamento); ou pode ser
uma manipulação que vise alterar os efeitos de outra manipulação (como nos
estudos clássicos de fisiologia do exercício).
De forma geral, quando a manipulação do comportamento é organizada de
forma a servir de medida para uma categoria de pesquisa, fala-se de modelos
comportamentais; estes são a redução de uma categoria de comportamentos 3 aos
seus elementos considerados, de forma teórica, como essenciais em um ambiente
padronizado, de forma a permitir o estudo das bases farmacológicas, biológicas ou
ambientais de cada uma destas categorias. Há diversos textos sobre modelos
(confira, por exemplo, Willner, 1991; Gouveia Jr, 1999). A leitura destes textos
pode ser feita sobre quem tiver mais curiosidade sobre este tema e por aqueles
que decidirem seguir a área.
Na escolha do comportamento a ser analisado, o conhecimento da ecologia
comportamental e da fisiologia da espécie é de suma importância. Tal fato evita a
antropomorfização dos estímulos e comportamentos – o famoso “Cânone de
Morgan” - ou seja, evita que o pesquisador interprete os dados observados
através de um tipo de psicologia ingênua (Folk psychology), baseado na leitura
3
Uma categoria de comportamentos inclui diversas classes de respostas, i.e., um grupo de coisas que o animal
faz com a mesma função e suas respectivas topografias, ou seja, a forma como isto é feito. Por exemplo, uma
categoria comportamental “agressão” implica nas classes de resposta de luta, fuga, observação, etc, e estas
podem incluir diversas formas de ocorrência, como displays e lutas físicas.
18
das reações do animal a partir do pressuposto de que elas correspondem a suas
próprias reações (o caso típico é do sujeito que considera que um peixe de fundo
– “limpa vidro” - isolado em um saco de plástico e exposto a luz nada de um lado
para o outro por estar “feliz”, e não como parte de um comportamento defensivo
de tentar buscar uma toca ou sombra). Exemplificamos: conhecer o que uma
espécie enxerga pode evitar o uso de estímulos “não visíveis” para estes animais.
Um modelo animal largamente utilizado é o Gymnotus sp (Carapó ou tuvira), um
peixe fracamente elétrico que usa um campo eletromagnético para se localizar no
ambiente. Uma leitura antropomórfica do experimento evitaria o uso deste
comportamento, com prejuízos claros para o conhecimento.
A construção de um modelo comportamental depende do embricamento
dos conhecimentos relativos aos comportamentos da espécie, das leis
comportamentais e da biologia, integrando o conhecimento naturalístico ao
laboratorial, de forma a subsidiar a interpretação de dados com vistas ao
desenvolvimento de tecnologias comportamentais médicas, industriais e de
pesquisa. Não se trata de tarefa fácil, e implica em uma leitura de causalidade
“aninhada” (“nested”) dos diversos níveis de organização do comportamento e das
diversas causas (Willner, 1991; Killeen, 2001; Barker et al., 2003)
19
Cap. 3
Modelos comportamentais animais com peixes
(Piscinos)
Abaixo iniciaremos a analise da bibliografia selecionada, tendo por
parâmetros 1) a disponibilidade desta; 2) a tentativa de incluir o máximo de
diversidade de modelos (de forma que não foram citados todos os trabalhos de um
único autor, quando a sua metodologia é igual); e 3) a inserção somente do que
constituía observação direta do ato comportamental, seja pelo observador, seja
por instrumentos, retirando da amostra estudos como de alimentação, mas que ao
invés de observarem o animal se alimentado, fizeram análise de conteúdos
estomacais, ou de reprodução que somente contaram filhotes vivos.
Uma análise da revisão citada por nós indica, inicialmente, uma dispersão
dos artigos consultados em diversas revistas de diversas áreas de conhecimento.
A tabela 3 apresenta as revistas que aparecem na nossa revisão. Pode-se notar
que há revistas de psicologia, biologia, toxicologia, zoologia e bioquímica, entre
outras. Tal fato nos indica uma larga dispersão dos dados, usos e objetivos das
pesquisas em comportamento, que se estende desde o estudo da recepção de
sons até efeitos de compostos químicos.
Esta dispersão dos dados, embora impeça que se acompanhe uma ou duas
revistas para dar-nos um panorama da área, exigindo buscas através de
ferramentas de busca e similares, indica uma ampla possibilidade de usos além da
20
biologia de peixes ou do estudo restrito de animais aquáticos. Nesta revisão,
encontramos trabalhos desde a década de 40 e é curioso notar que há um
deslocamento dos títulos, de revistas de psicologia e zoologia no inicio do século,
para revistas específicas de animais ou ambientes aquáticos e para revistas de
áreas de pouca relação direta com estas, o que indica dois movimentos diversos:
por um lado, a criação e/ou consolidação de uma área de estudo; por outro,
generalização dos modelos com objetivos de medida de outras variáveis.
Tabela 3 – Revisão de revistas (não exaustiva) onde foram encontraram artigos
sobre comportamento de peixes.
•
American Zoologist
•
Animal Behavior
•
Animal Learning and Behavior
•
Aquatic Toxicology
•
Behavioral and Neural Biology
•
Behavioral Brain Research
•
Behavioral Neurosciences
•
Behavioral Processes
•
Biosystems
•
Brain Research Bulletin
•
Brazilian J. of Biological and Medical Research
•
Bulletin of the Psychonomic Society
•
Comparative Biochemistry and Physiology
•
Ekolojija
•
Environmental Toxicology and Chemistry
•
Ethology
•
Hearing Research
•
Hormones & Behavior
•
J of Comparative Psychology
•
J of Experimental Biology
•
J. of Experimental Marine Biology and Ecology
•
J. of Experimental Biology
•
J. of Fish Biology
•
J. of General Physiology
•
J. of Genetic Psychology
•
Learning and Motivation
•
Neurobiology of Learning & Memory
•
Neurosciences Letters
•
Neurotoxycology and Teratology
•
Perceptual & Motor Skill
•
Physiology and Behavior
•
PNAS
•
Progress in Neuro-psychopharmacology & Biological
•
Psychological Records
Psychiatry
•
Psychological Reports
•
Psychonomic Sciences
•
The Sciences of Total Environment
•
Vision Research
•
Zebrafish
•
21
Vamos examinar cada um dos experimentos selecionados em relação a qual
tipo de comportamento é utilizado, segundo uma classificação prévia. Por vezes,
um mesmo experimento pode ser utilizado como modelo de mais de um tipo de
comportamento. Em cada sessão teremos uma tabela com o nome dos autores,
ano, objetivo original do trabalho, tipo de resposta medida e aparato. Quando um
tipo de manipulação couber em mais de uma categoria, será citada na tabela.
Inicialmente, vamos analisar o tipo de resposta utilizada em cada um dos
modelos selecionados, categorizada em atividade espontânea, quando a resposta
do animal não tem que ser ensinada ou treinada; respondente, quando uma
resposta já existente é relacionada por um treino ou manipulação a um estímulo
outro; operante, quando o controle da resposta é posterior à resposta do animal; e
misto, quando mais de um destes padrões estão colocados 4 .
Modelos que usam atividade espontânea
A Tabela 4 apresenta os trabalhos selecionados como utilizando atividade
espontânea, como variável principal.
Pode-se perceber que o animal mais usado é o goldfish (Carassius
auratus), seguido do Danio rerio (Paulistinha, zebrafish), sendo os experimentos
com o zebrafish os mais recentes – os experimentos mais antigos utilizam as
outras espécies. Tal aparecimento de artigos com o Danio rerio pode refletir a
4
Obviamente estas categorias podem ser questionadas, e um leitor com uma leitura mais etológica pode
reclamar dos termos utilizados, enquanto outro, com uma formação mais behaviorista, do uso de
comportamentos mistos (que seriam operantes) e da atividade espontânea, que seria um operante não
ensinado. Assumimos o uso “flácido” da teoria e não desconsideramos estas e outras críticas, mas avaliamos
que para a população alvo deste texto estas sutilezas teóricas não seriam o objetivo, ao menos neste momento.
22
disseminação do uso do Zebrafish como modelo biológico de desenvolvimento e
bioindicador (confira Henk et al, 2004, para uma análise da política do National
Institute of Health americano – NIH – com relação ao zebrafish e sua
disseminação), e estes dados seriam secundários a outras pesquisas biológicas
desenvolvidas.
Tabela 4 – Artigos selecionados que utilizam atividade espontânea como variável
dependente (resposta medida), conforme autor, ano, periódico e animal
experimental.
autor
ano
periodico
CRAWSHAW 1975 Comparative Biochemistry and Biology
especie
Ictalurus nebulosus
Lepomis macrochirus
KLEREKOPER, TIMMS, WESTLAKE, DAVY, MALAR & ANDERSON 1970
Animal Behavior
Carassius auratus
Acta Biotheoretica
Carassius auratus
NEVITT & HALL 1977
Perceptual & Motor Skills
Carassius auratus
PLAUT & GORDON 1994
J. of Experimental Biology
Danio rerio
Physiology and Behavior
Carassius auratus
Behavioral Brain Research
Carassius auratus
Physiology and Behavior
Danio rerio
Animal Behavior
Poecilia reticulata
MATTIOLI, NELSON, HUSTON & SPIELER 1998
Brain Research Bulletin
Carassius auratus
MOK & MUNRO 1998
Physiology and Behavior
Oreochromis niloticus
MATIS, KREERELOPER & GENSLER 1974
IIGO & TABATA 1996
SALAS, BROGLIO, RODRIGUEZ, LOPEZ PORTAVELLA & TORRES 1996
HURD,DEBRUYNE, STRAUME & CAHILL 1998
LACHLAN, CROOKS & LALAND 1998
Brazilian J. of Biological and Medical
Research
Danio rerio
BUDICK & O'MALLEY 2000
J. of Experimental Biology
Danio rerio
HERNANDEZ 2000
J. of Experimental Biology
Danio rerio
SERRA, MEDALHA & MATTIOLI 1999
Mas para que tipo de objetivo é utilizado a atividade espontânea? A tabela 5
sumariza os principais objetivos dos artigos selecionados.
23
Tabela 5 – objetivos e principais resultados segundo autor e ano dos artigos
selecionados na categoria atividade espontânea.
autor
CRAWSHAW
ano
Objetivo
Resultado
1975
Atividade espontânea
Alterações T dependentes
KLEREKOPER, TIMMS,
WESTLAKE, DAVY, MALAR &
ANDERSON
Teste de equipamento de monitoramento de A locomoção é sistemática
1970
MATIS, KREERELOPER &
nado
Testar padrões de oscilação do nado
A locomoção muda ao longo do tempo
GENSLER
1974
espontâneo
NEVITT & HALL
1977
Relação entre atividade, peso e temperatura Correlação parcial entre as variáveis
Medidas de metabolismo de nado entre
PLAUT & GORDON
1994
Sem diferenças
clones e selvagens
Determinar ciclo cronobiológico de atividade Os animais apresentam ciclo circadiano de atividade
IIGO & TABATA
1996
SALAS, BROGLIO, RODIRGUEZ,
LOPEZ PORTAVELLA & TORRES
1996
MATTIOLI, NELSON, HUSTON &
SPIELER
motora
Aprendizagem espontânea com dica em
Alterações relativas a constância, mas não as dicas em
labirinto em animais lesados
animais com ablação
Preferência condicionada por predisposição - Alteração de T dose dependente
1998
efeito de antagonista H1
1998
Ciclo circadiano de atividade
Variação circadiana da atividade
Discriminar variáveis no forrageamento
A preferência por seguir segue uma interação entre as
grupal
variáveis
HURD,DEBRUYNE, STRAUME &
CAHILL
LACHLAN, CROOKS & LALAND
1998
Efeitos de apomorfina e ablação sobre nado Padrão de atividade similar ao de mamíferos e ação das
MOK & MUNRO
1998
SERRA, MEDALHA & MATTIOLI
1999
BUDICK & O'MALLEY
manipulações
Preferência por escuridão
Preferência por escuridão
Etograma de nado
Variações intra-sujeitos e entre sujeitos, com repertórios
diferentes de nado em cada situação.
2000
Efeitos da morfologia da mandíbula sobre
HERNANDEZ
2000
comportamento alimentar
Dependente da morfologia
A tabela 5 dá uma idéia das possibilidades de uso de modelos
comportamentais animais piscinos, como o desenvolvimento de equipamentos,
24
fatores cronobiológicos, uso como modelos de estudo farmacológico, para
determinar diferenças entre metabolismo de espécies e efeitos de manipulações
neurológicas. Mas quais tipos de atividade e aparato ocorrem?
Na tabela 6, podemos ver o tipo de aparato utilizado para os estudos de
atividade espontânea e o tipo de atividade que é realizada. Uma breve olhada
indica que as atividades espontâneas mais comumente utilizadas são a
ambulação e a permanência, seguida de perto pela ingestão. Apenas poucos
autores separam as topografias, isto é, a forma como ocorre o nado, para
classificá-las.
Os aparatos mais usados são aquários sem outras modificações, sendo
que poucos automatizam as respostas e há modificações na cor do aquário e
apenas uma ocorrência de modificação da forma do aquário, com um labirinto em
X, ocorre.
No exame do método, vemos que a ambulação é feita contando números
de quadrantes percorridos ou o numero de cruzamentos entre ambientes do
aparato, como é o caso do labirinto em X e das caixas branco/preto. É curioso
notar que o uso de células fotoelétricas ocorre em trabalhos com Goldfish; tal uso
deve ser evitado, pois estes animais têm visão de ultravioleta, utilizado nas células
fotoelétricas, o que pode ser uma variável interveniente sobre os testes. O
trabalho de Budick e O’Malley (2000) usa placas de Petri; dado que o estudo
utiliza-se de larvas, esse equipamento se justifica como um aquário.
25
Tabela 6 – Artigos selecionados de atividade espontânea, com variável
dependente (medida) e descrição sumária do aparato utilizado.
Autor
ano
CRAWSHAW 1975
variáveis
Aparato
Nado
Aquario 3 l
Ingestão
Ventilação
KLEREKOPER, TIMMS, WESTLAKE, DAVY, MALAR &
Ambulação
Aquário 5x5x5 com células fotoelétricas
MATIS, KREERELOPER & GENSLER 1974
Ambulação
Aquário 5x5x5 com células fotoelétricas
NEVITT & HALL 1977
Ambulação
Aquário com sensores
Velocidade de nado
Aquário 30 l
Ambulação
Aquário com sensores
Permanência
Labirinto em X
ANDERSON 1970
PLAUT & GORDON 1994
IIGO & TABATA 1996
SALAS, BROGLIO, RODIRGUEZ, LOPEZ PORTAVELLA
& TORRES 1996
MATTIOLI, NELSON, HUSTON & SPIELER 1998
Ambulação
Permanência
Caixa branco/preto
Ambulação
HURD,DEBRUYNE, STRAUME & CAHILL 1998
LACHLAN, CROOKS & LALAND 1998
MOK & MUNRO 1998
SERRA, MEDALHA & MATTIOLI 1999
BUDICK & O'MALEY 2000
Atividade motora
Aquário
Forrageamento
Aquário colorido 91 x81 x30 cm
Ambulação
Campo aberto (Aquário redondo)
Tempo e ambulação
Caixa branco/preta
Nado de torno
Placas de Petri com Paramencium
Nado de fuga
Alimentação
HERNANDEZ 2000
Alimentação (tempo e
quantidade)
Aquário
26
Modelos que usam comportamentos que podem ser alvo de aprendizagem:
operantes, respondentes e mistos
Para os nossos propósitos, a aprendizagem pode ser definida como uma
modificação consistente de uma certa resposta (o que o organismo faz) ao longo
do tempo por força de uma variável controlável (Flaherty, 1985).
A aprendizagem é uma característica essencial de todo ser vivo e permite a
sua adaptação no meio. A aprendizagem pode ser definida em dois momentos: 1)
aquisição (ou aprendizagem em alguns teóricos) – o momento em que um
comportamento passa a ocorrer e se modifica ao longo do tempo; e 2)
manutenção (ou memória): quando a taxa de ocorrência de um comportamento
em uma dada situação é constante ou retoma níveis similares ao de suas ultimas
exposições quando da re-exposição 5 .
Classicamente, divide-se a aprendizagem entre aprendizagem respondente
ou pavloviana, quando um determinado comportamento já existente fica sob
controle de um estimulo neutro antecedente após um treino; e operante ou
instrumental, quando uma resposta fica sob controle de uma conseqüência desta
após um treino. As relações entre tipos de aprendizagem são um capitulo a parte
e também fonte de exaustivas controvérsias (para uma revisão, ver Flaherty, 1985,
ou Catania, 1999).
5
O termo memória é alvo de estudos e controversisas diversas. Para uma revisão, sugerimos Xavier (1993) e
Isquerdo (2002).
27
Manipulações que declaram usar cognição 6 normalmente tratam de
seqüências de estímulos que sinalizam a resposta em configurações variadas e
são exemplos de relações operantes/respondentes; aqui, estes estão classificados
como modelos mistos. As tabelas 7, 8 e 9 apresentam os artigos selecionados.
A análise das revistas em que são veiculados os artigos selecionados indica
uma forte presença de revistas de comportamento, só recentemente havendo uma
maior dispersão por revistas de biologia, toxicologia e outras áreas.
Nos comportamentos operante e respondente podemos observar a forte
presença do Goldfish (Carassius auratus), especialmente nos artigos mais antigos.
Já naqueles considerados como mistos há maior diversidade de espécies, com
destaque para o Danio rerio, que também se destaca nas tabelas anteriores como
animal experimental, especialmente nos artigos mais recentes.
6
Outra fonte de confusão, a palavra cognição. Para uma revisão veja Sternberg (2000).
28
Tabela 7 – Alguns artigos que usam comportamentos operantes, conforme autor,
ano e animal experimental utilizado
Autor
Ano
Periódico
PADILLA, PADILLA, KETTERER & GIACALONE 1970 Psychonomic Sciences
BEHREND & BITTERMAN 1963 JEAB
SCOBIE & HERMAN 1972 Learning and Motivation
DILL 1974 Animal Behavior
SCOBIE & GOLD 1975 Animal Learning and Behavior
WOLACH, MCHALE, BERSCHOT & FLEMING 1975 Psychological Reports
BRONSTEIN 1984
J of Comparative Psychology
COUVILLON 1984 J of Comparative Psychology
BROWN, SMITH & PETERS 1985 Psychological Reports
SHISHIMI 1985 J. of Comparative Psychology
DOUGLAS, EVA & GUTTRIDGE 1988 Behavioral Brain Research
BUDAEV 1997 J. of Comparative Psychology
Espécie
Carassius auratus
Carassius auratus
Carassius auratus
Danio rerio
Carassius auratus
Carassius auratus
Betta splendens
Carassius auratus
carassius auratus
Carassius auratus
Carassius auratus
Poecilia reticulata
MATTIOLI, SANTANGELO, COSTA &
VASCONCELOS 1997 Behavioral Brain Research
DÖOR & NIUMEYER 1997 Vision Research
MIZUKAMI, GUNJI & MIGITA 1999 Biosystems
Carassius auratus
Carassius auratus
carassius auratus
SPIELER, NELSON, HUSTON & MATTIOLI 1999 Neurosciences Letters
Carassius auratus
TALTON, HIGA &STADDON 1999 Behavioral Processes
carassius auratus
Progress in Neuro-psychopharmacology
MEDALHA, COELHO & MATTIOLI 2000 & Biological Psychiatry
Carassius auratus
LOPEZ, BINGMAN, RODRIGUEZ, GOMEZ &
SALAS 2000 Behavioral Neurosciences
SANTANGELO, MORATO & MATTIOLI 2001 Neurosciences Letters
WILLIAMS, WHITE & MESSER JR 2002 Behavioral Processes
Carassius auratus
Carassius auratus
Danio rerio
Notemigonus
WEBBER & HAINES 2003 Environmental Toxicology and Chemistry crysoleuca
CRAFT, VELKEY II, SZALDA-PETREE 2003 Behavioral Processes
Betta splendens
HIGA & SIMM 2004 Behavioral Processes
Betta splendens
COLWILL, RAYMOND. FERREIRA &
ESCUDERO 2005 Behavioral Processes
BILOTTA, RISNER, DAVIS & HAGGBLOOM 2005 Zebrafish
Danio rerio
Danio rerio
29
Tabela 8 – Alguns artigos que usam aprendizagem respondente, conforme
autor, ano e animal experimental utilizado
autor
ano
SCOBIE & HERMAN 1972
PRIEDE 1974
Periódico
Espécie
Learning and Motivation
J. of Experimental Biology
Carassius auratus
Salmo gairdineri
Carassius auratus/ salmo
ERSPAMER E MEYER 1978
Bulletin of the Psychonomic Society
BREUNING & WOLACH 1979
Psychological Record
Carassius auratus
ERSPAMER & MEYER 1979
Bulletin of the Psychonomic Society
Carassius auratus
WOLACH 1980
Psychological Record
Carassius auratus
ZELLA & HOLACH 1980
Psychological Record
Carassius auratus
gairdneri
BREUNING, FERGUNSON, MCHALEN &
MELVIN, PRENTICE-DUNN, ADANS &
HERRING III 1986
LAMING & MCKINNEY 1990
HALPERIN, DUNHAN & YE 1992
BRONSTEIN 1994
HALL & SUBOSKI 1995
J. of Comparative Psychology
Behavioral Neurosciences
Betta splendens
Carassius auratus
Behavioral Processes
Betta splendens
J-of-Comparative Psychology
Betta splendens
Neurobiology of Learning & Memory
Danio rerio
Comparative Biochemistry and
MANTEIFEL & KARELINA 1996
FAY 1998
DARLAD & DOWLING 2001
Physiology
Carassius auratus
Hearing Research
Carassius auratus
PNAS
Danio rerio
Neurotoxycology and Teratology
Danio rerio
LEVIN, CHRYSANTHIS, YACISIN &
LINNEY 2003
Brachyhypopomus
STODDARD, MARKHAM & SALAZAR 2003
J. of Experimental Biology
pinnicaudatus
BARRETO & VOLPATO 2004
Behavioral Processes
Oreochromis niloticus
MOREIRA & VOLPATO 2004
J of Fish Biology
Oreochromis niloticus
MOREIRA, PULLMAN & POTTINGER
2004
Hormones & Behavior
Oncorhicus mykiss
O Betta splendens teve uma redescoberta nos últimos anos, devido ao
interesse no estudo da agressão, comportamento do qual é um modelo clássico.
30
Por ser um peixe relativamente robusto (chega a 15 cm) é tão adequado para
manipulações estereotáxicas como o Carassius aurautus (possuindo, como este,
atlas adequado: Marino-Netto e Sabbatini, 1988), além de exigir poucos cuidados,
a espécie têm sido bastante utilizada. As espécies do gênero Oreochromis
apresentam maior intensidade de ocorrência nos últimos anos na literatura, devido
ao seu uso como animal de corte.
Tabela 9 - Alguns artigos que usam aprendizagem mista, conforme autor, ano e
animal experimental utilizado.
Autor
Ano
COLE & CALDWELL
1956
Periódico
J Comparative Physiology &
Psychology
SHAKLEE
LAMING & SAVAGE
JAKOBSEN & JOHNSEN
GERLAI & HOGAN
CAPURRO, REYES-PARADA,
OLAZABAL, PERRONE, SILVEIRA &
MACADAR
1963
1980
1989
1992
J. of Genetic Psychology
Behavioral and Neural Biology
Ethology
J. of comparative Psychology
1997
Comparative Biochemistry and
Physiology
NEMETH
1997
SMITH & WEIS
ANSTIS, HUTAHAJAN & CAVANAGH
FJELD, HAUGEN & VOLLESTAD
KRAUSE, HARTMANN & PRITCHARD
MIKLOSI, ANDREW & SAVAGE
1997
1998
1998
1998
1998
WEIS & WEIS
LOPES-CORREA & HOFFMANN
FUIMAN, SMITH & MALLEY
HSU & WOLF
MIKLÓSI & ANDREW
GERLAI, LAHAV, GOU & ROSENTHAL
PLAUT
SAMSON, GOODRIDGE, OLOBATUYI &
WEIS
PETRAUSKIENE
1998
1999
1999
1999
1999
2000
2000
J. of Experimental Biology
J. of Experimental Marine Biology and
Ecology
Vision Research
The Sciences of Total Environment
Animal Behaviour
Physiology and Behavior
J of Experimental Marine Biology and
Ecology
Physiology & Behavior
J. of Fish Biology
Animal Behavior
Behavioral Brain Research
????
J of Experimental Biology
2001
2002
Aquatic Toxicology
Ecology
Espécie
Carassius auratus
Cichlassoma meeki
Xyphophorus variatus
Danio rerio
Carassius auratus
Carassius auratus
Danio rerio
Macropodus opecularis
Gymnotus carapo
Hexagrammus
decagrammus
Fundulus heterocritus
Poecilia reticulata
Thymallus Thymallus
Danio rerio
Danio rerio
Fundulus heterocritus
Gymnotus carapo
Sciaenops ocellatus
Rivulus marmuratus
Danio rerio
Danio rerio
Danio rerio
Danio rerio
Oncorhicus mykiss
Seguindo o mesmo caminho feito anterior, analisaremos inicialmente o tipo
de objetivo e resultado e posteriormente qual tipo de resposta comportamental e
aparato de foi utilizado em cada uma das categorias apresentadas.
31
A tabela 10 apresenta os objetivos e resultados dos experimentos que
utilizaram respostas operantes.
Tabela 10 - Artigos selecionados de atividade operante, com objetivo e descrição
sumaria dos resultados obtidos
Autor
Ano Objetivos
BEHREND & BITTERMAN 1963 Esquiva de sidman
PADILLA, PADILLA, KETTERE &
GIACALONE 1970 Desamparo aprendido
SCOBIE & HERMAN 1972 Teste de choques para uso posterior
Efeitos da apresentação de modelos,
DILL 1974 filmagem de predador sobe fuga
DILL 1974 Efeitos da experiência sobre a esquiva
Resultado
obtido
Constata desamparo similar a mamíferos
Variação paramétrica da resposta e habituação
Variação entre apresentações
Habituação
SCOBIE & GOLD 1975 Efeito de reforçamento de baixas taxas
Aprendizagem similar a outras espécies
WOLACH, MCHALEN, BERSCHOT &
FLEMING 1976 Efeito de aquisição e extinção sucessiva Peixes se comportam como ratos
BRONSTEIN 1984 Uso de agressão como operante
Animais trabalham para agredir
COUVILLON 1984 Reforçamento diferencial em FI
Aprendizagem
BROWN, SMOTH & PETERS 1985 Desamparo aprendido
SHISHIMI 1985 Inibição latente
Desamparo aprendido
Existe inibição latente no goldfish
DOLGLAS, EVA & GUTTRIDGE 1988 Percepção de constância de tamanho
Constância de tamanho relatada
Padrões de personalidade, ambulação e
BUDAEV 1997 esquiva de predador
Relação entre padrões de exploração e esquiva
MATTIOLI, SANTANGELO, COSTA &
Efeito de Substância P em memória
VASCONCELOS 1997 apetitiva
Facilitação de memória por substancia p
DÖOR & NIUMEYER 1997 Aprendizagem de contraste de cores
DORR & NEUMEYER 1997 Contraste de cores
MIZUKA,I, GUNGI & MIJITA 1999 Discriminação simples
SPIELER, NELSON, HUSTON &
Efeitos de bloqueador H1 sobre
MATTIOLII 1999 aprendizagem e memória
TALTON, HIGA &STADDON 1999 VI e FI
Esquiva inibitória e efeitos de
MEDALHA, COELHO & MATIOLLII 2000 histaminérgicos
LOPEZ, BINGMAN, RODRIGUEZ,
Efeitos da ablação telencefálica sobre
GOMEZ & SALAS 2000 aprendizagem de labirinto
Esquiva inibitória - efeitos da substância
SANTANGELO, MORATO & MATTIOLII 2001 P
WILLIANS, WHITE & MESSER JR 2002 Resposta apetitiva na shuttlebox
Efeitos de dieta mercurial sobre esquiva
de presa
WEBBER & HAINES 2003 Predação
CRAFT, VELKEY II, SZALDA-PETREE 2003 Condicionamento de escolha
Condicionamento temporal (FI) passar
HIGA & SIMM 2004 em uma argola
COLWILL, RAYMOND. FERREIRA &
ESCUDERO 2004 Discriminação de cores
BILOTTA, RISNER, DAVIS &
HAGGBLOOM 2005 Discriminação apetivitiva
Determinação do espectro de visão
Ocorrência de contraste de cores
O animal aprende nesse esquema
Diferenças entre animais tratados e controle
O animal aprende nesses esquemas
Modulação histaminérgica sobre comportamento
Dissociação de dicas e lugar se mantém no animal
lesado
Efeitos da substância P facilita a aprendizagem
O peixe aprendeu
Alterações dose-dependentes da predação
O peixe apresenta aprendizagem de escolha
O peixe apresenta aprendizagem de controle temporal
Aprendizagem e extinção em todas as situações
O peixe aprende discriminação simples
32
A observação dos trabalhos apresentados indica uma ampla utilização dos
modelos
operantes
para
diversas
respostas
comportamentais,
algumas
caracteristicamente alvo de estudos operantes clássicos ligados aos efeitos de
história comportamental sobre a aprendizagem: desamparo aprendido, efeitos de
experiência, extinções e aquisição.
Outros estudos que tentam mostrar a ocorrência de fenômenos já
demonstrados em mamíferos, como ratos, entram nesta categoria: o efeito de
reforçamento de baixas taxas, reforçamento diferencial em intervalo fixo, inibição
latente, estudos em intervalo variado e fixo (VI e FI), resposta apetitiva na
shuttlebox, condicionamento de escolha, condicionamento temporal.
Alguns trabalhos se dedicam a estudos mais paramétricos, com o estudo de
testes de choques, o uso de agressão como operante, entre outros, por fim,
alguns trabalhos utilizam operantes para demonstrar experimentalmente efeitos de
drogas e toxinas, manipulações cirúrgicas, ou medir limiares de percepção ou
gerar padrões correlacionais (de personalidade). De maneira geral, embora o
numero de espécies seja pequeno, todos os trabalhos relatam sucesso em suas
manipulações.
O exame da tabela seguinte (Tabela 11) indica qual operante foi utilizado
em cada estudo e qual o aparato. Uma breve análise desta tabela indica que as
respostas motoras utilizadas podem ser divididas em 3 grandes tipos: nadar para
fora ou dentro de algum compartimento, permanecer (ou seja, não nadar) e
apertar um disco ou uma chave, algo parecido com uma resposta de focinhar em
ratos ou bicar em pombos, ou caçar/ agredir. Desta forma, em que difere estas
manipulações daquelas colocadas como de operante livre?
33
Em primeiro lugar, elas estão relacionadas a um arranjo de estímulos
externos diversos; por outro lado, exige uma modificação consistente deste
comportamento relacionada à apresentação do estímulo escolhido, ou seja,
aprendizagem.
Esta necessidade se expressa na maior variabilidade de aparatos
utilizados, que embora sejam aquários, apresentam adaptações de forma, como
nos labirintos; de outros componentes colocados para serem manipulados pelos
animais, como argolas, barras, chaves ou luzes; ou o uso de aparelhagens
simples baseadas em aversão ou comportamento espontânea como elemento de
aprendizagem, como na caixa branco e preto ou nas modificações da eficácia de
caça.
O uso de operantes apresenta uma certa constância ao longo dos anos
vistos, no entanto, a linguagem utilizada modifica-se, sendo abandonado um
vocabulário típico do behaviorismo, uma linha teórica da psicologia, por um
vocabulário mais descritivo dos objetivos do estudo.
Consideramos tal fato vantajoso, dado que permite a uma gama de
profissionais maior trabalharem juntos, e diminui tolas discussões sobre que tipo
de vocabulário é o mais adequado pra falar sobre comportamento.
34
Tabela 11 - Artigos selecionados de atividade operante, com variável dependente
(medida) e descrição sumaria do aparato utilizado
Autor
Ano
Variáveis
BEHREND & BITTERMAN
PADILLA, PADILLA, KETTERE &
GIACALONE
1963 Cruzar ambiente para postergar choque
1970 Fuga (nado)
Shuttle box
SCOBIE & HERMAN
1972 Fuga (nado)
Shuttle box
DILL
1974 Fuga (nado)
Aquários adaptados
SCOBIE & GOLD
WOLACH, MCHALEN, BERSCHOT &
FLEMING
1975 Operante livre (nado), tempo sem resposta
Caixa de Skinner
Operante livre (nado), com extinções ou não inter1976 periodos
Aquário de 8 compartimentos relacionados
BRONSTEIN
1984 Resposta em FI (chave) para acesso a espelho
Aquário/caixa de resposta motora
COUVILLON
1984 Respostas em uma chave
Aquário com luzes e alvos (chaves)
BROWN, SMOTH & PETERS
1985 Esquiva de duas vias (nadar) de choque
Shuttle box
SHISHIMI
1985 Atividade/taxa de resposta (nadar/chave)
Activimentro/caixa de Skinner/ Shuttle box
DOLGLAS, EVA & GUTTRIDGE
1988 Escolha (nadar)
BUDAEV
MATTIOLI, SANTANGELO, COSTA &
VASCONCELOS
1997 Ambulação (campo aberto), esquiva de predador
Labirinto com buracos
Campo aberto hexagonal 0.9m/labirinto de
inspeção de predador/
DÖOR & NIUMEYER
1997 Manutenção de aprendizagem (permanência)
Resposta operante de matching de cores/
Resposta operante sobe disco de luz (focinhar e
1997 nadar)
MIZUKA,I, GUNGI & MIJITA
1999 Taxa de respostas certas treinadas
SPIELER, NELSON, HUSTON & MATTIOLI 1999 Escolha
TALTON, HIGA &STADDON
1999 Chave
Aparato
shuttlebox
Labirinto em Y branco/preto
Aquário adaptado com projeção de luzes
Aquário de 3 compartimentos
Caixa de Skinner
MEDALHA, COELHO & MATTIOLI
2000 Esquiva de ambiente claro (nado)
LOPEZ, BINGMAN, RODRIGUEZ, GOMEZ
& SALAS
2000 Resposta operante em labirinto (nado)
Caixa branco/preta
SANTANGELO, MORATO & MATTIOLI
2001 Latência de saída (nado)
Caixa branco/preta
WILLIANS, WHITE & MESSER JR
2002 Alternação (nado)
Shuttle box
WEBBER & HAINES
2003 Esquiva/caça de camarões
Aquário e filmadora
CRAFT, VELKEY II, SZALDA-PETREE
2003 Operante de resposta de entrada(nado)
Passagem por argola de tubos em função do
2004 tempo
Labirinto
Argola de tubos/espelho
2004 Escolha (permanência)
Labirinto em T
HIGA & SIMM
COLWILL, RAYMOND. FERREIRA &
ESCUDERO
BILOTTA, RISNER, DAVIS & HAGGBLOOM 2005 Operante de resposta de entrada (nado)
Labirinto em X com dicas
Labirinto
A Tabela 12 apresenta os dados relativos aos testes aqui classificados de
respondentes, ou seja, aqueles que usam um comportamento já observado em um
animal, mas tentam colocar esta resposta sob controle de um estimulo
35
antecedente, ou buscar correlatos fisiológicos do qual este comportamento possa
vir a ser uma medida indireta.
Podemos perceber que as respostas cardíacas e respiratórias parecem ser
as preferidas como forma de estudo, embora respostas apetitivas e agressivas
também possam ser utilizadas, ou sons. A principal característica aqui é um
deslocamento da aprendizagem para estudos de percepção ou emoção, nos quais
as respostas incondicionadas têm papel essencial.
Também aqui, os trabalhos visam os correlatos comportamentais de
alterações fisiológicas com maior freqüência, de certa forma tentando usar a
metodologia comportamental como uma forma de “entrar” na fisiologia do animal.
Tal fato se reflete no uso de respostas como variável dependente, com mais
respostas como display, preferência (que, embora tenha um elemento motor
operante, é, em geral, inata), resposta de aglutinação, forma de onda elétrica em
peixes fracamente elétricos, mas ainda com várias respostas de ambulação, ou de
exploração, como a resposta orptomotora e de permanência.
Os aparatos utilizados vão refletir estas escolhas dos pesquisadores, com
geradores de estímulos como ondas sonoras e luzes, e aparelhagem especial,
como um aparelho especial de condicionamento de resposta ventilatória e
frequencímetros.
36
Tabela 12 - Artigos selecionados de atividade respondente, com objetivo e
descrição sumária dos resultados obtidos
Autor
Ano
SCOBIE & HERMAN
PRIED
ERSPAMER E MEYER
BREUNING & WOLACH
ERSPAMER
BREUNING, FERGUNSON,
MCHALEN & WOLACH
ZELLA & HOLACH
MELVIN, PRENTICE-DUNN, ADANS
& HERRING III
LAMING & MCKINNEY
HLAPERIN, DUNHAN & YE
BRONSTEIN
HALL & SUBOSKI
MANTEIFEL & KARELINA
FAY
DARLAD & DOULING
LEVIN, CHRYSSANTHIS, YACISIN &
LINNEY
STODDARD, MARKHAM & SALAZAR
BARRETO & VOLPATO
MOREIRA, PULLMAN& POTTINGER
MOREIRA & VOLPATO
Objetivos
1972 Teste de choques para uso posterior
Efeitos do nado em taxa cardíaca de peixes
1974 vagotimizados
1978 Condicionamento de resposta cardíaca
Contraste negativo (alteração US-CS) em função da
1979 forma de alojamento
Resultado
Variação da resposta em função da intensidade
e salinidade
Relação clara ente velocidade e taxa cardíaca
Ausência de condicionamento
Tipo de alojamento muda curva de resposta
1979 Condicionamento de resposta cardíaca
Condicionamento
Condicionamento de resposta respiratória - efeitos de
1980 magnitude do choque
A resposta é modulável pela magnitude
1980 Condicionamento de contraste (pavloviano)
Os peixes aprendem
1986 Efeitos da punição sobre agressão
Efeito da ablação sobre habituação de resposta
1990 cardíaca
Diminuição do display em função da punição
1992 Efeitos de privação social sobre agressão
1994 Fatores paramétricos na agressão
Testar aprendizagem respondente da reação de
1995 alarme
Ablação não altera habituação
Maior agressão em isolados
Fatores de predictibilidade, habituação e
sensibilização podem ser observados.
Aprendizagem Ocorre
1996 Testar aversão condicionada a alimento
Aprendizagem de aversão
1998 Percepção de sons
O peixe reconhece tons de forma mamífera
2001 Dependência de cocaína
Cocaína altera os três testes
2003 Efeito de pesticida
Efeitos de diminuição da aprendizagem
2003 Efeitos de serotoninérgicos sobre resposta elétrica
Correlação entre resposta ventilatória e indicadores
2004 de estresse
Seleção de resposta para estresse, condicionamento
2004 e extinção pavloviana.
A onda é modulada
A resposta ventilatória é indicador de estresse
crônico, mas não agudo
O animal aprende
2004 Condicionamento pavloviano de resposta a estressor Condicionamento
37
Tabela 13 - Artigos selecionados de atividade respondente, com variável
dependente (medida) e descrição sumaria do aparato utilizado
Autor
Ano
SCOBIE & HERMAN
PRIED
ERSPAMER E MEYER
Variáveis
Aparato
1972 Reação motora (levantamento da nadadeira dorsal)
Shuttlebox
1974 Taxa cardíaca e velocidade
Aquário em O
1978 Taxas em relação a US
Aquário de contenção/ frequencímetro
Aquários isolados com caixas acústicas e
1979 Atividade (ambulação) entre US-CS
luz
1979 Taxas em relação a US
Aquário de contenção/ frequencímetro
MCHALEN & WOLACH
1980 Parada de comportamento ventilatório
Condicionador de resposta respiratória
ZELLA & HOLACH
1980 Parada de comportamento ventilatório
Condicionador de resposta respiratória
1986 Duração do display
Aquário e gerador de choques
1990 Taxa de resposta cardíaca e ventilação
Luzes p/ sobressalto
HLAPERIN, DUNHAN & YE
1992 Display
Aquários/espelho
BRONSTEIN
1994 Display
Aquários
1995 Reação de alarme (nado e aglutinação)
Aquário
1996 Ingestão
Aquário 1 l
1998 Freqüência respiratória
Condicionador de resposta respiratória
BREUNING & WOLACH
ERSPAMER
BREUNING, FERGUNSON,
MELVIN, PRENTICE-DUNN, ADANS
& HERRING III
LAMING & MCKINNEY
HALL & SUBOSKI
MANTEIFEL & KARELINA
FAY
Aquário com dicas visuais/ T-maze/resposta
DARLAD & DOULING
2001 Preferência de lugar
orptomotora
2003 Treino de preferência lateral condicionada
Aquário de escolha com três câmeras
2003 Forma de onda
Frequencímetro
2004 Indicadores fisiológicos e resposta ventilatória
Observação em aquário
2004
Tanque/luz
2004 Cortisol plasmático
Luz e confinamento
LEVIN, CHRYSSANTHIS, YACISIN
& LINNEY
STODDARD, MARKHAM &
SALAZAR
BARRETO & VOLPATO
MOREIRA, PULLMAN&
POTTINGER
MOREIRA & VOLPATO
38
A Tabela 14 apresenta os artigos classificados como usando atividade
mista. Estes trabalhos são em suas maioria paramétricos, ou seja, exploram os
efeitos de dimensões da variável independente sobre a resposta comportamental
dos sujeitos. Seus objetivos são bastante variáveis, com motivação, lateralização,
busca de estratégias e correlatos com tetrápodes aparecendo com bastante
constância.
Tanto metodologias operantes como respondentes estão presentes, por
vezes, dentro de manipulações de variáveis respondentes com vistas a sinalizar
ou alterar variáveis operantes. Tais relações ficam claras na tabela 15, em que se
observam os tipos de variáveis dependentes mensuradas.
Podemos perceber que estas variáveis são padrões mais complexos de
comportamento que se articulam com variações ambientais. Os aparatos
utilizados, por sua vez, primam pela simplicidade, utilizado variáveis naturalísticas
com pouca ou nenhuma variação.
39
Tabela 14 - Artigos selecionados de atividade mista, com objetivo e descrição
sumaria dos resultados obtidos.
Autor
Ano
COLE & CALDWELL
SHAKLEE
LAMING & SAVAGE
JAKOBSEN & JOHNSEN
Objetivos
Resultado
1955 Uso de luz para motivar exploração
A luz diminui tempo e aumenta acertos
1963 Medidas de medo
Diferenças inter-espécies
Mapeamento de alterações leva a concluir que
estas seguem um padrão mamífero
1980 Alterações fisiológicas no alerta e fuga
GERLAI & HOGAN
CAPURRO, REYES-PARADA,
OLAZABAL, PERRONE, SILVEIRA &
MACADAR
1992 Interação inato aprendido na agressão
Efeito concorrente do feromônio
Os animais aprendem a agredir imitando a
outros
Esquiva e agressão de descarga elétrica / efeitos
1997 NMDA
NMDA aumenta agressão
NEMETH
1997 Tipo de captura de presa / segundo tipo de presa
Diferenças entre estratégias
SMITH & WEIS
MIKLOSI, ANDREW & SAVAGE
KRAUSE, HARTMANN &
PRITCHARD
FJELD, HAUGEN & VOLLESTAD
WEIS & WEIS
ANSTIS, HUTAHAJAN &
CAVANAGH
CORREA & HOFFMANN
MIKLÓSI & ANDREW
FUIMAN, SMITH & MALLEY
HSU & WOOF
GERLAI, LAHAV, GOU &
ROSENTHAL
PLAUT
SAMSON, GOODRIDGE,
OLOBATUYI & WEIS
PETRAUSKIENE
1989 Efeitos de feromônio de alarme sobre alimentação
1997 Efeitos da poluição sobre comportamento de predação Alterações relativas à taxa de poluição
1998 Verificar lateralização
Escolha de companheiros em função de estado
1998 nutricional
Efeito do metil-mercurio sobre a ingestão alimentar
1998
1998 Efeito do chumbo sobre predação, nado e atividade.
Lateralização tipo tetrapode
Escolha de co-especifico/maior escolha de
nutrido/melhor forrageamento destas duplas
Diminuição dose-dependente da eficácia
alimentar
Alterações dose dependentes em todos os
parâmetros
Animal Apresenta resposta
1998 Resposta orptomotora
Variação das descargas elétricas em função de drogas
1999 em teste de novidade
Alterações dose e droga dependente
1999 Lateralização e decisão de comer
Testar padrões de sobressalto e variável de controle,
1999 se acústica ou visual.
Efeitos da experiência de vencer/perder sobre o
1999 comportamento
Há lateralização da decisão de morder
A resposta é mais controlada pela visão que
pela acústica
Diferenças no comportamento segundo a
história de vida
2000 Efeitos de etanol em parâmetros comportamentais
Etanol altera todos os parâmetros
Testar efeito de diferença de tamanho da nadadeira na
2000 velocidade de nado
Diferenças relativas a tamanho da nadadeira
2001 Efeitos do MeHg
Alterações dose dependentes
2002 Agressão após exposição a metais pesados
Alterações pela exposição
40
Tabela 15 - Artigos selecionados de atividade mista, com variável dependente
(medida) e descrição sumaria do aparato utilizado.
autor
Ano
COLE & CALDWELL
SHAKLEE
LAMING & SAVAGE
JAKOBSEN & JOHNSEN
GERLAI & HOGAN
CAPURRO, REYES-PARADA,
OLAZABAL, PERRONE, SILVEIRA &
MACADAR
NEMETH
SMITH & WEIS
MIKLOSI, ANDREW & SAVAGE
KRAUSE, HARTMANN &
PRITCHARD
FJELD, HAUGEN & VOLLESTAD
WEIS & WEIS
ANSTIS, HUTAHAJAN &
CAVANAGH
CORREA & HOFFMANN
MIKLÓSI & ANDREW
FUIMAN, SMITH & MALLEY
HSU & WOOF
GERLAI, LAHAV, GOU &
ROSENTHAL
PLAUT
SAMSON, GOODRIDGE,
OLOBATUYI & WEIS
PETRAUSKIENE
Variáveis
1955 Tempo, ambulação e erros
Aparato
Labirinto de oito braços,
complexo.
1963 Ambulação e esquiva de luz
Aquário com luzes/campo aberto
1980 Alterações fisiológicas a apresentação de estímulos
Aquário/ fisiógrafos
1989 Densidade de presas e snacks
Aquário 26 x 10 x 10 cm.
1992 Diversos parâmetros
Aquário
1997 Taxas de disparo
Osciloscópio
1997 Tipo de captura/movimentos
Filmagem
1997 Snack sobre camarões
Aquário e camarões
Espelhos/objetos em aquário de
1998 Perseguição a espelho e aproximação de objeto novo 20/40/20 cm
1998 Outros espécimes/privação
Labirinto
1998 Aquários e dafnias
Alterações dose dependentes
1998 Snacks em artemia/ nado/ ambulação
1998 Resposta orptomotora
Aquário e artemias
Aquário esférico com estímulos
circulatórios
1999 Descargas elétricas
Osciloscópio
1999 Morder
Aquário com estímulos laterais
1999 Espaço percorrido/velocidade
Tanque 150 l
1999 Luta/resultados
Agressão/ambulação/preferência por
grupos/comportamento antipredatorio/preferência por
2000 escuridão
Aquário
Resposta a espelho/schooling/
caixa claro-escuro/ modelo de
predador
2000 Ambulação e nado de performance (induzido)
Tubo de nado forçado
2001 Nado/captura de presas
Aquário
2002 Etograma agressão/atividade total
Aquário
Modelo de quê?
O objetivo declarado de nosso percurso até aqui foi demonstrar alguns
modelos experimentais para peixes; no entanto, o que é um modelo
comportamental?
Conforme definimos anteriormente, para cientistas do comportamento,
“modelo” não é uma espécie especifica, mas um set up, ou seja, um arranjo
composto por um aparato e um procedimento que caracteriza uma manipulação
41
relativa a uma categoria de comportamento. Desta forma, o mesmo aparato pode
servir para modelos diferentes: por exemplo, a caixa branco/preto tanto pode ser
utilizada como modelo para esquiva inibitória como para preferência; no primeiro
caso, uma porta e posta em seu meio, o animal em sua parte escura e a latência,
(isto é, o tempo que o animal demora a sair) do lado escuro é computado. Este
procedimento é repetido algumas vezes com intervalo entre as tentativas, e o
resultado é uma curva de aprendizagem, com o aumento das latências ao longo
das tentativas.
Na outra manipulação, a preferência, há duas portas corrediças no centro
que delimitam um compartimento inicial; após uma breve habituação, as portas
são abertas e o tempo despendido em cada ambiente, taxa de cruzamentos e o
número de acessos a cada ambiente são computados.
As categorias de comportamento que os modelos buscam replicar visam o
entendimento de patologias e comportamento não patológicos típicos de nossa
espécie, buscando homologias e analogias que dependem de opções teóricas
subsidiadas pelas teorias psicológicas, e pelo conhecimento de neurociências e de
elementos de fisiologia comparada.
Desta forma, comportamentos aparentemente distantes de uma patologia
podem ser utilizados para o estudo desta. Por exemplo, um teste feito em ratos, o
teste de enterramento, mede a velocidade de um roedor para se enterrar em uma
cama de maravalha, após uma modificação no ambiente. Considerado um teste
de ansiedade, pois responde a fármacos ansioliticos, possui uma semelhança no
mínimo tênue com os comportamentos de ansiedade em humanos.
42
Para que um modelo animal se torne útil, além de apresentar um set up
bem descrito e uma consistência teórica que o habilite, deve apresentar, em graus
variados, uma validade farmacológica, ou seja, uma resposta a drogas próxima à
patologia ou o grupo de patologias a que o modelo se propõe mimetizar; uma
validade de face, ou seja, uma semelhança na apresentação do comportamento
que facilite seu reconhecimento e interpretação; e uma validade preditiva, ou seja,
que este modelo permita o desenvolvimento de fármacos e tratamentos.
Tomamos a liberdade de classificar a bibliografia aqui apresentada dentro
de possíveis modelos (Tabela 16). Tal classificação não visa ser uma forma
absoluta ou taxativa, dado que muitos destes modelos não tem uma validação
farmacológica ou sua fisiologia bem estudada, mas tentamos gerar uma possível
agenda de pesquisa para os grupos que estejam estabelecidos nas áreas de
modelos, indicando possíveis caminhos e com o estimulo para que mais
pesquisadores motivem-se a militar nesta área.
A classificação foi feita com base na analogia de modelos em ratos (por
exemplo, o uso da inibição latente como modelo de esquizofrenia) ou elementos
naturalísticos da resposta do animal (todos os modelos que envolvem a
apresentação de um estressor e medida de fuga foram classificados como
possíveis modelos de ansiedade/medo).
43
Tabela 16 – distribuição dos autores citados segundo possível uso dos
experimentos como modelos.
Condicionamento clássico: BRONSTEIN,1994
Agressão: MELVIN, PRENTICE-DUNN, ADANS & HERRING III,1986; HLAPERIN, DUNHAN & YE ,1992; GERLAI & HOGAN,
1992 BRONSTEIN,1994; NEMETH, 1997; GERLAI, LAHAV, GOU & ROSENTHAL, 2000
Ansiedade/ medo: BREUNING, FERGUNSON, MCHALEN & WOLACH,1980; BUDAEV,1997; CAPURRO, REYES-PARADA,
OLAZABAL, PERRONE, SILVEIRA & MACADAR,1997; CORREA & HOFFMANN,1999; DILL,1974; ERSPAMER,1979; FUIMAN,
SMITH & MALLEY,1999; GERLAI, LAHAV, GOU & ROSENTHAL,2000; HALL & SUBOSKI,1995; HSU & WOOF,1999; JAKOBSEN &
JOHNSEN,1989; KLEREKOPER, TIMMS, WESTLAKE, DAVY, MALAR & ANDERSON,1970; LAMING & MCKINNEY,1990;
MATTIOLI, NELSON, HUSTON & SPIELER,1998; MEDALHA, COELHO & MATTIOLI,2000; MOK & MUNRO,1998; SANTANGELO,
MORATO & MATTIOLI,2001; SCOBIE & HERMAN,1972; SERRA, MEDALHA & MATTIOLI,1999; SHAKLEE,1963; WEBBER &
HAINES,2003; ZELLA & HOLACH,1980; BEHREND & BITTERMAN, 1963
Aprendizagem/ cognição: SCOBIE & GOLD ,1975; LEVIN, CHRYSSANTHIS, YACISIN & LINNEY,2003; MATTIOLI, NELSON,
HUSTON & SPIELER,1998; MIZUKA,I, GUNGI & MIJITA,1999; DOLGLAS, EVA & GUTTRIDGE,1988; WOLACH, MCHALEN,
BERSCHOT & FLEMING,1976; MANTEIFEL & KARELINA,1996; BREUNING & WOLACH,1979; MIZUKA,I, GUNGI & MIJITA,1999;
BRONSTEIN,1994,GERLAI & HOGAN,1992; BRONSTEIN,1984; WILLIANS, WHITE & MESSER JR,2002; SPIELER, NELSON,
HUSTON & MATTIOLI,1999
Cronobiologia: IIGO & TABATA,1996; HURD,DEBRUYNE, STRAUME & CAHILL,1998; MATIS, KREERELOPER &
GENSLER,1974
Dependência: DARLAD & DOULING, 2001
Depressão: PADILLA, PADILLA, KETTERE & GIACALONE,1970; BROWN, SMOTH & PETERS, 1985
Desenvolvimento de comportamento complexo: BUDICK & O'MALEY, 2000
Doença cardíaca: PRIED, 1974
Escolha social?: KRAUSE, HARTMANN & PRITCHARD, 1998
Esforço físico: PLAUT & GORDON,1994; PLAUT, 2000
Esquizofrenia: SHISHIMI,1985
Estresse: BARRETO & VOLPATO,2004; MOREIRA, PULLMAN& POTTINGER,2004 ERSPAMER E MEYER, 1978; MOREIRA &
VOLPATO,2004; CRAWSHAW, ?
Exploração: CORREA & HOFFMANN,1999
Lateralidade: MIKLOSI, ANDREW & SAVAGE,1998; MIKLÓSI & ANDREW, 1999
Memória: KLEREKOPER, TIMMS, WESTLAKE, DAVY, MALAR & ANDERSON,1970; SCOBIE & HERMAN,1972; SCOBIE &
HERMAN,1972; WILLIANS, WHITE & MESSER JR,2002; DARLAD & DOULING,2001; MIZUKA,I, GUNGI & MIJITA,1999; SPIELER,
NELSON, HUSTON & MATTIOLI,1999; MEDALHA, COELHO & MATTIOLI,2000; SANTANGELO, MORATO & MATTIOLI,2001;
TALTON, HIGA &STADDON,1999; DOLGLAS, EVA & GUTTRIDGE,1988; MATTIOLI, SANTANGELO, COSTA &
VASCONCELOS,1997; BILOTTA, RISNER, DAVIS & HAGGBLOOM,2005; SALAS, BROGLIO, RODIRGUEZ, LOPEZ PORTAVELLA
& TORRES,1996; COLE & CALDWELL,1955; LOPEZ, BINGMAN, RODRIGUEZ, GOMEZ & SALAS,2000; CRAFT, VELKEY II,
SZALDA-PETREE,2003; HIGA & SIMM,2004; COLWILL, RAYMOND. FERREIRA & ESCUDERO,2004
Screening de drogas: SAMSON, GOODRIDGE, OLOBATUYI & WEIS,2001; STODDARD, MARKHAM & SALAZAR,2003
Surdez: FAY, 1998
Temporização do comportamento: TALTON, HIGA &STADDON, 1999
Toxicologia: PETRAUSKIENE,2002; FJELD, HAUGEN & VOLLESTAD, 1998; SMITH & WEIS, 1997; WEIS & WEIS, 1998
Visão: DARLAD & DOULING,2001; ANSTIS, HUTAHAJAN & CAVANAGH,1998; DORR & NEUMEYER,1997; DOLGLAS, EVA &
GUTTRIDGE,1988; COLWILL, RAYMOND. FERREIRA & ESCUDERO,2004
44
Uma análise do conteúdo apresentado na tabela 16 indica que o uso de
peixes pode ser tão amplo quanto o de roedores, e envolve aspectos diversos do
comportamento, que podem ser utilizados para estudo da aprendizagem (tanto em
condicionamento clássico, como respondente), cognição, memória, aspectos
desenvolvimentais do comportamento, percepção e reação ao tempo e a variáveis
cronobiológicas, além de aplicações nos estudos de psicopatologia, toxicologia,
aspectos complexos do ponto de vista da neurologia e de doenças cardíacas. De
certa forma, quase tudo o que tradicionalmente fazemos com roedores pode ser
feito com peixes, exceto, talvez, privá-los de água.
45
Capitulo 4
Modelos de ansiedade piscina
Nesta sessão examinaremos especificamente os trabalhos de nossa
seleção que versam sobre ansiedade/medo e estresse. Para tanto, iniciaremos
falando sobre como ansiedade, medo e estresse se relacionam, suas definições e
sobre a forte relação homológica dos sistemas monoaminérgicos que medeiam a
ansiedade em peixes e mamíferos.
Em situações em que um animal é predado, ou se vê defronte de um perigo
intenso, uma seqüência previsível de reações ocorre, com mecanismos de
deflagração, topografia e fisiologia correlata extremamente similares em todos os
mamíferos (Blanchard, Yudko, Rodgers & Blanchard, 1993).
A reação de defesa é uma seqüência de 3 níveis (Graeff, 1994; Blanchard
& Blanchard, 1988), relacionada com 4 estratégias comportamentais: imobilização
(freezing), fuga, agressão defensiva ou submissão (Zangrossi Jr., 1996; Blanchard
e Blanchard, 1988).
Cada um destes níveis está relacionado com a probabilidade e
proximidade de um estímulo potencialmente aversivo. Estes comportamentos são
altamente plásticos em sua apresentação, podendo assumir topografias diversas
46
segundo a historia de vida. A Tabela 17 sumariza os comportamentos, emoções e
as áreas de ativação em mamíferos, onde este fenômeno está mais estudados.
Tabela 17 – Nível de defesa, substrato neural e emoção relacionada (adaptada de
Graeff, 1994).
Perigo
Comportamentos
Emoção
Sistema nervoso central
relacionada
Potencial Levantamento de risco
Distal
Congelamento,
Ansiedade
fuga, Medo
esquiva.
Proximal
Fuga, luta.
Amígdala, Sistema septo-hipocampal (SHS)
Matéria cinzenta periaquiductal-dorsal (PAG), Núcleos
medianos da rafe; SHS, hipotálamo medial
pânico
PAG
Tal tipo de comportamento implica em uma base aminérgica conhecida
desde a década de 80 (Gray,1982) e que é mais recentemente relacionada com a
ação da serotonina (5-HT), que é o principal neurotransmissor das regiões citadas
e que tem, segundo alguns autores, um sistema dual de ativação de todos os
comportamentos em função da proximidade do estimulo (McNaughton & Corr,
2004).
Tal grupo de comportamentos também está correlacionado a relação de
decisão sobre a estratégia a ser tomada (esquiva ou luta defensiva), evento este,
sumarizado nas Figura 1 e relacionado a áreas do encéfalo e síndromes clinicas
na figura 2.
47
Figura 1 - processos envolvidos na reação de defesa (Graeff, 1993)
48
Figura 2 – papel dual da serotonina sobre a ativação de áreas neurais e
comportamento em função da proximidade do estimulo e da decisão de fuga
defensiva ou esquiva defensiva, segundo hierarquia de ativação (de baixo para
cima), nível neural e complexidade do comportamento ativado, relacionado com
síndrome comportamental clinica relacionada. (onde: TOC: Transtorno Obsessivocompulsivo; TAG : Transtorno de Ansiedade Generalizada; 5-Ht: % hidroxitripitamina (serotonina); NA: Noradrenalina.. Observe que quanto maior a distância
defensiva, isto é, a distância do estimulo aversivo, mais complexa e menos motora
a reação. Adaptado de McNaughton & Corr, 2004
A complexidade do comportamento descrito indica que diferentes modelos
devem estar relacionados a diferentes síndromes e áreas do cérebro, mas que
compartilham uma base bioquímica e anatômica comuns, relacionada a serotonina
(File, 1996; Stanford, 1996).
49
Tal base é conservada em todos os vertebrados, não sendo diferente nos
peixes (confira, por exemplo, Ritchie, Livingston, Hughes, McAdoo & Leonard,
1989; Lundin & Holmgren, 1989). Esta manutenção evolutiva permite que modelos
de ansiedade e depressão possam ser consistentemente generalizados entre
espécies, desde que conhecida a estrutura da ecologia comportamental de cada
uma delas e sua base bioquímica. Não estamos dizendo com isto, que as
espécies são iguais entre si, mas que o conhecimento produzido sobre uma
espécie pode colaborar com o conhecimento produzido sobre outras, nesses
aspectos evolutivamente conservados.
Podemos ver que toda a teoria por trás do entendimento da ansiedade
implica em uma relação com uma possível punição ambiental. Tal relação não
ocorre somente no sistema nervoso central, mas implica em alterações hormonais
não especificas, chamadas comumente de estresse, com ênfase nas alterações
do
eixo
hipotalâmico-hipofisário-adrenal
e
simpático-adreno-medular
em
vertebrados, relativas à necessidade de manutenção de homeostase do
organismo e acompanhada de alterações comportamentais e fisiológicas.
Por um mecanismo de generalização, o estresse passou a designar uma
variedade de manipulações de variáveis externas (os estressores), que também
passaram
a
ser
chamados
de
estresse,
bem
como
suas
respostas
comportamentais relacionadas. Desta forma, diversos modelos que aqui
discutiremos como modelos de ansiedade são tratados como modelos de estresse
na literatura, e buscam a identificação de correlatos comportamentais das
alterações fisiológicas (Volpato & Fernandes, 1994). Na área de estudos de
comportamento de peixes, tais modelos são, comumente voltados para a
50
piscicultura e o crescimento de animais de corte. A Tabela 18 é uma tentativa de
classificação dos modelos aqui apresentados.
Numa primeira aproximação, podemos acreditar que todos os níveis de
apresentação da ansiedade estão representados, mas a classificação como feita
por nós não está livre de problemas. Vários dos modelos apresentados são
cotidianamente utilizados em roedores; no entanto, em roedores estes modelos
costumam apresentar formas mistas de ansiedade, que envolvem os dois ou três
níveis. Cremos que estes problemas devem se manter entre peixes.
Tabela 18 – Sumula dos modelos apresentados neste texto. A letra R indica
similar em roedores.
Aparato/modelos
Exploração de ambiente novo R
Preferência por escuridão (caixa branco/preta) R
Esquiva de luz R
Condicionamento de resposta cardíaca (?)
Condicionamento de resposta ao estressor(?)
Condicionamento de resposta respiratória (?)
Efeito de feromonios sobre alimentação
Variação de descargas elétricas de peixe fracamente elétrico
em função de manipulações ambientais
Sobressalto R
Esquiva inibitória R
Esquiva de duas vias R
Esquiva de Sidman R
Efeitos de luta sobre comportamento posterior
Resposta ventilatória ao estresse R
A seguir tentamos de classificação dos modelos de ansiedade aqui
apresentados.
Numa primeira aproximação, podemos acreditar que todos os
níveis de apresentação da ansiedade estão representados, mas a classificação
como feita por nós não está livre de problemas. Vários dos modelos apresentados
51
são cotidianamente utilizados em roedores; no entanto, em roedores estes
modelos costumam apresentar formas mistas de ansiedade, que envolvem os dois
ou três níveis (tanto tipo-medo, como tipo-ansiedade) (conf. Zangrossi Jr, 1996).
Cremos que estes problemas devem se manter entre peixes.
Os modelos já utilizados com roedores podem ser mais facilmente
validados no futuro, já que a sua comparação pode ser feita de forma mais direta.
Estes estão assinalados na tabela.
A tabela 19 apresenta um rearranjo das tabelas anteriores, indicando o tipo
ansiedade provavelmente apresentada nos artigos citados e seu método, bem
como aparato. Podemos perceber que a resposta dos possíveis modelos
utilizados pode ser dividida em modelos que exigem aprendizagem e que não
exigem esta; destes, podemos falar de modelos que exigem aprendizagem
respondente e aqueles que usam aprendizagem operante.
Do que conhecemos do modelo de ansiedade, podemos considerar alguns
modelos como sendo de ansiedade (aqueles que o perigo é potencial), de medo
(onde o perigo é distal, na forma de um predador ou punição) e de pânico (onde o
perigo é proximal). Por economia, colocamos o único modelo que se aproximaria
desta forma de apresentação junto com os modelos de medo.
Em nosso laboratório, 4 modelos estão em uso: a preferência claro escuro,
a esquiva inibitória de estímulo naturalístico, a esquiva de duas vias de estímulo
naturalístico e o labirinto em Y. Consideramos que os 3 primeiros são potenciais
modelos de ansiedade e o último, de aprendizagem operante e memória.
52
Tabela 19 – classificação dos possíveis modelos de ansiedade piscina
segundo exigência de aprendizagem e técnica.
Ansiedade
Modelos que exigem aprendizagem
tecnica:
Condicionamento de preferência
Dissonância por aprendizagem
concorrente (supressão?)
Esquiva ativa
Esquiva de Sidman
Esquiva inibitória
aparato
Caixa branco/preto
Aquário 26 x 10 x 10 cm.
AUTOR E ANO
MATTIOLI, NELSON, HUSTON & SPIELER,1998
JAKOBSEN & JOHNSEN, 1989
Aquários adaptados
shuttlebox
Caixa branco/preta
DILL,1974
BEHREND & BITTERMAN,1963
MEDALHA, COELHO & MATTIOLI,2000;
SANTANGELO, MORATO & MATTIOLI,2001
Modelos de atividade espontânea
Modelos de confronto
Modelo de:
Confronto agonistivo
labirinto de inspeção de predador/
Aquário e filmadora
Modelo de predador
AUTOR E ANO
CAPURRO, REYES-PARADA, OLAZABAL,
PERRONE, SILVEIRA & MACADAR, 1997
BUDAEV, 1997
WEBBER & HAINES, 2003
GERLAI, LAHAV, GOU & ROSENTHAL, 2000
aparato
Campo aberto (Aquário redondo)
AUTOR E ANO
MOK & MUNRO,1998
Campo aberto (quadrado)
CRAWSHAW,1975; HSU & WOOF,1999
Campo aberto hexagonal 0.9m/
Campo aberto (quadrado)
BUDAEV,1997
KLEREKOPER, TIMMS, WESTLAKE, DAVY, MALAR
& ANDERSON,1970
GERLAI, LAHAV, GOU & ROSENTHAL,2000
SERRA, MEDALHA & MATTIOLI,1999;
GERLAI, LAHAV, GOU & ROSENTHAL,2000
CORREA & HOFFMANN,1999
aparato
Osciloscópio
Esquiva de predador
Modelos de exploração
Modelo de:
Exploração
Resposta a espelho
Caixa branco/preta
Teste de novidade
Osciloscópio
Medo
Modelos que exigem aprendizagem
Modelo de:
Aprendizagem respondente
Fuga
aparato
Condicionador de resposta respiratória
Shuttlebox
Aquário com luzes/campo aberto
AUTOR
ZELLA & HOLACH,1980
SCOBIE & HERMAN,1972
SHAKLEE,1963
ano
Modelos de atividade espontânea
Modelos de confronto
Sobressalto
Tanque 150 l
FUIMAN, SMITH & MALLEY,1999
Estresse (correlato funcional de ansiedade crônica)
Modelo de:
Condicionamento de resposta
respiratória - respondente
Condicionamento de resposta
cardíaca - respondente
Condicionamento de resposta
cardíaca- respondente
aparato
Observação em aquário
Condicionador de resposta respiratória
Luz e confinamento
Tanque/luz
Aquário de contenção/ frequencímetro
Luzes p/ sobressalto
AUTOR
ano
BARRETO & VOLPATO,2004
BREUNING, FERGUNSON, MCHALEN &
WOLACH,1980
MOREIRA & VOLPATO,2004
MOREIRA, PULLMAN& POTTINGER,2004
ERSPAMER,1979; ERSPAMER E MEYER,1978
LAMING & MCKINNEY,1990; HALL &
SUBOSKI,1995
53
Dados de nosso laboratório.
Embora vejamos uma serie de potenciais modelos de ansiedade, em nosso
laboratório temos desenvolvido trabalhos com dois possíveis modelos de
ansiedade, por um lado, a esquiva inibitória, utilizando estimulo naturalístico, por
outro, a caixa branco-preto, para estudo de preferência. Claro escuro.
Preferência por escuridão ou claro escuro: O teste de preferência é realizado
em um aquário (15 x 10 x 45 cm) com dois compartimentos, um branco e outro
preto, com uma área central (5 cm) delimitada por portas corrediças e que é usado
como compartimento de inicio do teste. Após 5 minutos de habituação, as portas
são suavemente removidas, dando acesso a ambos os compartimentos,
permitindo ao animal explorar livremente o ambiente durante um período de 15
(quinze) minutos. , Registra-se principalmente o tempo despendido em cada
compartimento além do numero de sujeitos que preferem cada ambiente na
primeira latência, bem como a latência de saída do compartimento inicial para
qualquer um dos lados.
Figura 3 – Aparato para preferência branco-preto. Observe as portas corrediças.
Uma variação deste, com uma única porta pode ser utilizado para esquiva
inibitória do lado branco.
54
Os resultados de nosso laboratório coadinam com a literatura (Serra et al,
1999), e vêm indicando maior preferência pelo lado escuro (Gouveia et al, 2005b,
Gouveia et al, 2006, submetido), sendo a
padronização comportamental,
realizada a partir da utilização de aquários branco-branco e preto-preto em
contraste ao de duas cores., Esse modelo foi originalmente aplicado em Danio
(Danio rerio) por Serra et al (1999), sendo posteriormente testado em Goldfish
(Carassius auratus) (Gouveia et al, 2005), Lambari (Axtyanax autiparanae), Néon
(paracheirodon sp), Zebrafish (Danio rerio), Tuvira (Gymnotus sp), Lebiste (Poecila
reticulata) e Tilapia (Oreochromis niloticus).
Esquiva inibitória de estimulo naturalístico: É composta pelo mesmo aparato,
porém, sem portas, após 5 minutos de exploração livre, um dos lados é punido
pela queda de um peso metálico de 15 g preso a uma trave de uma altura de 22
cm do aquário, sempre do mesmo lado. A queda do peso é repetida cada vez que
o animal cruza a linha média do aquário, até o limite de 7 vezes ou quando o
animal fica mais de 300 s sem cruzar a linha. Espécies já testadas: Lambari
(Astyanax altiparanae), Goldfish (Carassius auratus), Zebrafish (Danio rerio)
(Anexo 1)
55
Figura 4 – Aparato para esquiva de uma via de estimulo naturalistico. Uma
variação deste, com uma única porta pode ser utilizado para esquiva inibitória do
lado branco
De forma proposital, não apresentamos nenhum trabalho de nosso
laboratório. Esta decisão teve duas motivações: por um lado, preservar os nossos
dados, já que em grande parte estes estão submetidos ou em fase final de
redação; e, por outro, permitir que descrevêssemos de forma detalhada os
modelos que vimos utilizando em nosso laboratório e que já foram eventualmente
comunicados em congressos.. Esperamos que tal descrição sirva de estímulo para
que outros pesquisadores venha a se engajar nesta área de modelos
comportamentais piscinos.
56
Quadro 2 – dicas para a construçãode uma shuttlebox com estimulo
naturalístico.
Descrição de materiais para Sutllebox:
- Aquário:
- dimensões: 45 cm comprimento x 15 cm altura x 10 cm largura;
- Barreira de acrílico:
- dimensões: 14,5 cm comprimento x 9,5 cm largura x 1 mm espessura;
- Placas laterais de acrílico - (02) duas:
- dimensões: 26 cm comprimento x 30 cm altura x 1 mm espessura;
- Suporte para parafusos:
- dimensões: 19 cm comprimento x 12,5 cm altura x 11,5 cm largura;
- 02 (dois) conjuntos: parafuso com quatro porcas, sendo que cada conjunto tem
espessura de 11 mm, comprimento de 19 cm e peso de 110 gramas;
- Base de compensado (madeira):
- dimensões: 54 cm comprimento x 1,5 cm altura x 33 cm largura;
- 04 (quatro) pés de madeira com 30 cm de altura cada;
- travessa frontal ligando dois pés frontais: 45,5 cm comprimento x 03 cm
altura x 1 cm espessura;
- 02 (dois) pregadores;
- 02 (duas) arruelas metálicas;
- 10 (dez) roldanas pequenas (cinco em cada suporte);
- Linha de polietileno preta 2,5 mm espessura x 2,5 m comprimento;
57
Referências
Anstis S., Hutahajan P. & Cavanagh P. (1998). Optomotor test for wavelength
sensitivity in guppyfish (Poecilia reticulata). Vision Research, 38, 1, 4553.
Barker G. A., Derr P. G., Thompson N. S. (2004). The perils of confusing nesting
with chaining in psychological explanations. Behavior and Philosophy,
32, 293-303.
Barreto R. E., Volpato G. L. (2004). Caution for using ventilatory frequency as an
indicator of stress in fish. Behavioral Processes, 66, 1, 43-51.
Behrend E.R. & Bitterman, M.E. (1963). Sidman avoidance in the fish. J. of
Experimental Analysis of Behavior, 6, 1, 46-62.
Bilotta, J., Risner, M.L., Davis, E.C. & Haggbloom, S.T. (2005) Assessing
Appetitive Choice Discrimination Learning in Zebrafish. Zebrafish, 2, 4,
259-268.
Blanchard, D. C. & Blanchard, R. J. (1990). An Ethoexperimental approach to
biology of emotions. Annual Review of Psychology, 39, 43-68.
Blanchard, R. J., Yudko, E. B., Rodgers, R. J., Blanchard, D. C. (1993). Defense
system
psychopharmacology:
an
ethological
approach
to
the
pharmacology of fear and anxiety. Behavioral Brain Research, 58, 1-2,
155-65.
58
Bolis C.L., Piccolella, M., Dalla Valle A.Z. & Rankin, J. (2001). Fish as model in
pharmacological and biological research. Pharmacological Research,
44, 4. 265-280
Breuning, S.E. & Wolach, A.H. (1979). Sucessive negative contrasts effects for
activity-condicioned goldfish (Carassius auratus) as a function of the
house condictions. The Psichological Record, 29, 245-254.
Breuning, S.E., Fergunson, D.G., McLahe, M.A. & Wolach, A.H. (1980). Shifts in
magnitude of a shock US for goldfish (Carassius auratus) in a respiratory
conditioning situation. The Psichological Record, 30, 367-375.
Bronstein, P.M. (1984). Agonistics and reproductive interactions in Betta
splendens. J of Comparative Psychology, 98, 421- 431.
Bronstein, P.M. (1994). On the predictability, sensibilization, and habituation of
aggression in males bettas (Betta splendens). J. of comparative
Psychology, 1994,108,45-57.
Brown, G.E., Smith, P.J. & Peters, R. B. (1985). Effects of escapable versus
inescapable shocks on avoidance behavior in the goldfish (Carassius
auratus). Psychological reports, 57, 1027-1030.
Budaev, S.V. (1997). “Personality” in the guppy (Poecilia reticulata): A
correlacional study of exploratory behavior and social tendency. J. of
Comparative Psychology, 111, 4, 399-311.
Budick, S.A. & O’Malley, D.M. (2000). Locomotor repertoire of the larval zebrafish:
swimming, turning and prey capture. Journal of Experimental Biology
203, 2565– 2579
59
Capurro, A., Pakdaman, K., Perrone, R. & Macadar, O. (1999). Analysis of the
jamming avoidance response in the electric fish Gymnotus carapo.
Biological Cybernetics, 80, 4, 269–283
Catania A.C. (1999). Aprendizagem - Comportamento, Linguagem e Cognição.
Porto Alegre: ArtMed
Cole, M. B. & Caldwell, W. E. (1956). The utilization of light as exteroceptive
motivation in the comet goldfish (Carassius auratus). J of Comparative
Physiology and Psychology, 49, 1, 71-76.
Colwill, R.M., Raymond, M.P., Ferreira, L. & Escudero, H. (2005) Visual
discrimination
learning
in
zebrafish
(Danio
rerio).
Behavioral
Processes;70, 19–31
Cosme Salas, C. B., Emilio-Durán, A. G., Ocaña, F. M., Jiménez-Moya, F., &
Rodríguez, F (2006) Neuropsychology of Learning and Memory in Teleost
Fish. Zebrafish,. 3, 2, 157-171
Cosme-Salas-Garcia, C. B., Rodriguez-Fernandez, F.,
Lopez-Garcia, J.C.,
Portavella-Garcia, M. & Ruiz, B.T. (1996) Telencephalic Ablation in
Goldfish Impairs Performance in a 'spatial Constancy' Problem But Not in
a Cued One. Behavioural Brain Research, 79, 1-2, 193-200.
Couvillon, P.A. (1984). Performance of goldfish (Carassius auratus) in patterned
sequences of rewarded and nonrewarded trials. Journal of Comparative
Psychology, 98, 3, 333-44
Craft, B. B., Velkey, A.J-II & Szalda-Petree, A. (2003). Instrumental conditioning of
choice behavior in male Siamese fighting fish (Betta splendens).
Behavioural Processes, 63, 171-175.
60
Crawshaw, L. I. (1975). Twenty-four hour records of body temperature and activity
in bluegill sunfish (Lepomis macrochirus) and brown bullheads (Ictalurus
nebulosus). Comparative Biochemistry and Physiology, 51A, 11-14.
Darland, T. & Dowling, J. E. (2001). Behavioral screening for cocaine sensitivity in
mutagenized zebrafish. PNAS, 98, 11691-11696.
Dill, L.M. (1974) The escape response of the Zebra Danio (Brachydanio rerio). I.
The stimulus for escape. Animal Behaviour 22, 711-722
Dill, L.M. (1974) The escape response of the Zebra Danio (Brachydanio rerio). II.
The effect of experience. Animal Behaviour 22, 723-730
Donahoe, J. W. & Palmer, D. C. (1994). Learning and complex behavior.
Boston, MA: Allyn & Bacon.
Döor, S. & Neumeyer, C. (1997). A simultaneous color contrast in goldfish – a
quantitative study. Vision research, 37, 12, 1581-1593.
Douglas, R.H., Eva, J.& Guttridge, N. (1988) Size constancy in goldfish (Carassius
auratus) Behavioral Brain Research, 30, 1, 37-42
Emde, G. (2000). Electroreception. In D. H. Evans (2000). Physiology of Fishes.
London. CRC Press.
Erspamer, R.T. & Meyer, M.E. (1974). Heart rate conditioning in Goldfish
(Carassius auratus) and no in rainbow trout (Salmo gairdneri). Bulletin of
the psychonomic society, 11, 6, 374-378.
Erspamer, R.T. & Meyer, M.E. (1979). Heart rate conditioning of Goldfish,
Carassius auratus, with intermittent vs. continuous CS. Bulletin of the
psychonomic society, 13, 6, 381-382
61
Fay, R.R. (1998) Perception of two-tone complexes by goldfish (Carassius
auratus). Hearing Research, 120, 17-24.
File, S. E. (1996).
Recent developments in anxiety, stress, and depression.
Pharmacology Biochemistry and Behavior,54, 1, 3-12.
Fjeld, E., Haugen, T.O., & Vollestad, L.A. (1998). Permanent impairment in the
feeding
behavior
of
grayling
(Thymallus
thymallus)
exposed
to
methylmercury during embryogenesis. Science of Total Environment,
213, 1-3, 247-54
Flaherty, C. F. (1985), Animal Learning and Cognition. Alfred A. Knopf, Inc. New
York.
Fuiman, L. A., Smith, M. E. & Malley, V. (1999). Ontogeny of the rotine swinming
speed and startle responses in red drum, with a comparison response to
acoustic and visual stimuli. J. of Fish Biology, 55, Sup A, 215-226.
Gerlai, R., Hogan, J. A. (1992). Learning to find the opponent: an ethological
analysis of the behavior of paradise fish (Macropodus opercularis) in intraand interspecific encounters. J of Comparative Psychology,106, 3, 30615.
Gerlai, R., Lahav, M., Guo, S. & Rosenthal, A. (2000). Drinks like a fish: zebra fish
(Danio rerio) as a behavior genetic model to study alcohol effects.
Pharmacology Biochemistry and Behavior, 67, 4, 773-82.
Gouveia Jr, A. (1999). Modelos animais em psicopatologia: Breves notas
introdutórias. Estudos de Psicologia, 16, 1, 13-16,
Gouveia JR, A., Zampieri, R. A., Ramos, L. A., Silva, E. F., Matiolli, R. & Morato,
S. (2005) Prefference of goldfish (Carassius aurautus)for dark places.
62
Revista de Etologia, 7, 2.
Graeff, F. G. (1994). Neuroanathomy and neurotransmitter regulation of defensive
behaviors and related emotions in mammals. Brazilian J. of Medical and
Biological Research, 27, 811-819.
Gray, J. (1982) The Neuropsychology of anxiety : an enquiry into the function
of the septo-hippocampal system. Oxford University Press New York.
Hall, D. & Suboski, M.D. (1995). Visual and olfatory stimuli in learned release of
alarm reaction by Zebra Danio Fish (Brachydanio rerio). Neurobiology of
Learning & Memory, 63, 3, 1095-9564.
Halperin, J.R.P., Dunham, D.W. & Ye, S. (1992). Social isolation increases social
display after priming in Betta splendens but decreases aggressive
readness. Behavioral Processes, 28, 13-32.
Hernandez, L.P. (2000). Intraspecific scaling of feeding mechanics in an
ontogenetic series of zebrafish, Danio rerio Journal of Experimental
Biology, 203, 19, 3033-3043.
Higa, J. J. & Simm, L. A. (2004).Interval timing in Siamese fighting fish (Betta
splendens) Behavioral Processes, 67, 3, 501-509.
Hsu, Y.;& Wolf, L. L. (1999). The winner and loser effect: integrating multiple
experiences. Animal Behaviour, 57, 4, 903-910.
Hurd M.W., Debruyne J., Straume M. & Cahill G.M. (1998) Circadian rhythms of
locomotor activity in zebrafish. Physiology & Behavior, 65, 3, 465-72.
Iigo, M & Tabata, M. (1996) Circadian rhythms of locomotor activity in the goldfish
Carassius auratus. Physiology & behavior 60, 33, 775-781.
Isquerdo, I. (2002) Memória. Porto Alegre. Artes médicas.
63
Jakobsen, J. & Johnsen, G. H. (1989) The influence of alarm substance on feeding
in zebra danio. fish ( Brachydanio rerio). Ethology 82, 325-7.
Killeen PR (2001). The four causes of behavior. Current Directions in
Psychological Sciences, 10, 136-140.
Kleerekoper, H, Timms, A.M., Westlake, G.F., Davy, F.B., Malar, T. & Anderson,
V.M. (1970). An analysis of locomotor behaviour of goldfish (Carassius
auratus). Animal Behaviour,18, 2, 317-30.
Krause, J., Hartmann, N. & Pritchard, V.L. (1998). The influence of nutritional state
on shoal choice in zebrafish, Danio rerio. Animal Behaviour, 4, 711-775.
Lachlan, R. F., Crooks, L. & Laland, K. N. (1998). Who follows whom? Shoaling
preferences and social learning of foraging information in guppies Animal
Behaviour, 56, 1, 181-190.
Laming, P. R. & Savage, G. E. (1980) Physiological changes observed in the
goldfish (Carassius auratus) during behavioral arousal and fright.
Behavioral and Neural Biology, 29, 2, 255-74.
Laming. P.R. & McKinney, S.J. (1990). Habituation in goldfish (Carassius auratus)
is impaired by increased interestimulus interval, interval variability and
telencephalic ablation. Behavioral Neurosciences, 104, 6, 869-875.
Lehner, P.N. (1996). Handbook of ethological methods (second ed.).
Cambridge University Press, Cambridge.
Levin E.D., Chrysanthis E., Yacisin K. & Linney E. (2003). Chlorpyrifos exposure of
developing zebrafish: effects on survival and long-term effects on
response latency and spatial discrimination. Neurotoxicology and
Teratology, 25, 1, 51-57.
64
Lopes-Correa, S. A. & Hoffmann, A. (1999). Effect of drugs that alter alertness and
emotionality on the novelty response of a weak electric fish, Gymnotus
carapo. Physiology & behavior, 65, 4-5, 863-869.
López J.C., Bingman V.P., Rodríguez F., Gómez Y. &
Salas C. (2000)
Dissociation of place and cue learning by telencephalic ablation in goldfish
Behavioural Processes, 25, 81-93
Lorenz, K. (1983) Ele Falava com os Mamíferos, as Aves e os Peixes. Lisboa:
Publicações Europa América
Lucas J.J. & Hen R. (1995). New players in the 5-HT receptor field: Genes and
knockouts. Trends in Pharmaceutical Sciences, 16, 246-252
Lundin, K. & Holmgren, S. (1989 )The occurrence and distribution of peptide- or 5HT-containing nerves in the swimbladder of four different species of
teleosts (Gadus morhua, Ctenolabrus rupestris, Anguilla anguilla, Salmo
gairdneri). Cell and Tissue Research ,257, 3, 641 - 647
Manteifel, Y.B. & Karelina, M.A. (1996). Conditioned food aversion in the goldfish,
Carassius auratus. Comparative Biochemistry and Physiology A, 115,
31–35.
Marino-Neto, J. & Sabbatini, R.M.E. (1988). A stereotaxic atlas for the
telencephalon of the Siamese fighting fish (Betta splendens). Brazilian J.
of Medical and Biological Research, 21, 971-986.
Matis,J.H., Childers,D.R. & Kleerekoper, H. (1974) A stochastic locomotor control
model for the goldfish (Carassius auratus), Acta Biotheoretica, 23, 45–54
65
Mattioli R., Nelson C.A., Huston J.P. & Spieler R.E. (1998). Conditioned placepreference analysis in the goldfish with the H1 histamine antagonist
chlorpheniramine. Brain Research Bulletin, 45, 1, 41-4.
Mattioli R., Santangelo E.M., Costa A..C & Vasconcelos, L. Substance P facilitates
memory in goldfish in an appetitively motivated learning task. Behavioral
Brain Research, 85, 1, 117-20
McNaughton, N. & Corr, P. J. (2004) A two-dimensional neuropsychology of
defense: fear/anxiety and defensive distance. Neuroscience and
Biobehavioral Reviews. 28, 285–305
Medalha C.C., Coelho J.L., Mattioli R. (2000). Analysis of the role of histamine in
inhibitory avoidance in goldfish Progress in Neuropsychopharmacology
Biological Psychiatry,24, 2, 295-305.
Melvin, K.B., Prentice-Dunn, S., Adams, C.H. & Herring III, G.H. (1986). Curvilinear
effects of severity of punishment on species-typical aggressive display in
Betta splendens. J. of comparative psychology, 100, 2, 188-193.
Miklosi A.& Andrew R.J. (1999). Right eye use associated with decision to bite in
zebrafish. Behavioral Brain Research, 105, 2, 199-205.
Miklosi, A., Andrew, R.J. & Savage H. (1998) Behavioural lateralisation of the
tetrapoid type in the zebrafish (Brachydanio rerio). Physiology &
Behavior, 63, 1,127-35
Mizukami, E. Gunji, Y.P. & Mijgta, M (1999) Learning process by goldfish and its
use of a local site as a map Biosystems,54,1-2, 91-104
66
Mok, E.Y. & Munro, A.D. (1998). Effects of dopaminergic drugs on locomotor
activity in teleost fish of the genus Oreochromis (Cichlidae): involvement of
the telencephalon. Physiology & Behavior, 64, 3, 227-34.
Moreira P.S., Pulman K.G. & Pottinger T.G. (2004). Extinction of a conditioned
response in rainbow trout selected for high or low responsiveness to
stress. Hormones & Behavior, 46, 4, 450-457.
Moreira, P. S. A. & Volpato, G. L. (2004) Conditioning of stress in Nile tilapia.
Journal of Fish Biology 64, 4, 961-969.
Nemeth, D. (1997). Modulation of attack behavior and its effect on feeding
performance in a trophic generalist fish. Journal of Experimental
Biology, 200, 15, 2155-2164.
Nevitt JR & Hall R (1977) Correlation of activity and weight in comet goldfish
(Carassius auratus) at differing water temperatures. Perceptual and
Motors Skills, 45, 1, 81-2
Padilla, A. M., Padilla, C., Ketterer, T. & Giacalone, D. (1970). Inescapable shocks
an subsequent escape avoidance conditioning in goldfish, carassius
auratus. Psychonomic Science, 20, 295-296.
Plaut, I. (2000). Effects of fin size on swimming performance, swimming behavior,
and routine activity of zebrafish, Danio rerio . J. of Experimental
Biology., 203,.4, 813-820.
Plaut, I., & Gordon, M.S. (1994). Swimming metabolism of cloned and wild type
zebra fish, Brachydanio rerio. J. of Experimental Biology, 194, 209-223
Pough, F.H., Janis, C.M. & Heiser, J.B. (Org.) (2003). A vida dos vertebrados. 3ª
Edição. S. Paulo. Atheneu Editora.
67
Priede, I.G. (1974). The effect of swimming activity and section of the vagus nerve
on heart rate in rainbow trout; Journal of Experimental Biology, 60, 305319.
Prut L & Belzung C. (2003). The open field as a paradigm to measure the effects of
drugs on anxiety like-behaviors: a review. European Journal of
Pharmacology, 463: 3-33.
Ritchie, T. C., Livingston, C. A., Hughes, M. G., McAdoo D. J. & Leonard, R. B.
(1989). The distribution of serotonin in the CNS of an elasmobranch fish:
immunocytochemical and biochemical studies in the Atlantic stingray,
Dasyatis sabina. J of Comparative Neurology, 221, 4, 429-43.
Samson, J.C., Goodridge, R., Olobatuyi, F. & Weis, J.S. (2001) Delayed effects of
embryonic exposure of zebrafish (Danio rerio) to methylmercury (MeHg).
Aquatic Toxicology, 51, 4, 369-76
Santangelo, E. M., Morato, S. & Mattioli, R.(2001). Facilitatory Effect of Substance
P on Learning and Memory in the Inhibitory Avoidance Test for Goldfish.
Neuroscience Letters, 303, 2, 137-139.
Scobie, S. R. & Herman, H.B. (1972). Detection, reaction, escape and debilitation
thresholds for electric shock in Goldfish. Learning and motivation, 3,
442-456.
Scobie, S.R., Gold, D.C. (1975) Differential reinforcement of low rates in goldfish.
Animal learning and behavior,3, 2, 143-146.
Serra E. L., Medalha C.C. & Mattioli R. (1999) Natural preference of zebrafish
(Danio rerio) for a dark environment. Brazilian J. of Medical and
Biological research, 32, 1551-53.
68
Shaklee, A. B. (1963). Comparative studies of temperament: fear responses in
different species of fish. J. of Genetic Psychology, 102, 295-310.
Shishimi, A. (1985) Latent inhibition experiments with goldfish (Carassius auratus).
Journal of Comparative Psychology, 99, 3, 316-27
Smith, G. M. & Weis, J. S. (1997). Predator-prey relationships in mummichogs
(Fundulus heteroclitus (L.)): Effects of living in a polluted environment.
Journal of Experimental Marine Biology and Ecology, 3, 209, 1, 75-87.
Spieler, R.E., Nelson, C.A., Huston, J.P. & Mattioli, R. (1999) Post-trial
administration of H-1 histamine receptor blocker improves appetitive
reversal
learning
and
memory
in
goldfish,
Carassius
auratus
Neurosciences Letters , 277, 1, 5 – 8.
Stanford, S.C. (1996). Stress: A major variable in the psychopharmacologic
response. Pharmacology Biochemistry and Behavior, 54, 1, 211-7.
Stenberg, R. J. (2000) Psicologia Cognitiva . Porto Alegre
Stoddard, P.K., Markham, M. R. & Salazar, V. L. (2003). Serotonin modulates the
electric waveform of the gymnotiform electric fish Brachyhypopomus
pinnicaudatus. J. of Experimental Biology, 206, 1353-1362.
Talton, L., Higa, J.J., & Staddon, J.E.R. (1999). Interval Schedule Performance in
the Goldfish (Carassius auratus). Behavioural Processes, 45, 193-206.
Thorndike, E. L. (1911). A note on the psychology of fishes. American Naturalist,
XXXIII,
396
(consultado
em
http://psychclassics.yorku.ca/Thorndike/Animal/fish.htm#f1 , ultimo acesso
em 10/07/2006).
69
Val, A. L. & Almeida-Val, V.M. (Org.) (1999). Biology of tropical fishes. Manaus.
Instituto Nacional de Pesquisa Amazônica.
Volpato, G. L. & Fernandes, M. O. (1994) Social control of grown in fish. Brazilian
J. of Medical and Biological research, 27, 797-810.
Webber H.M. & Haines T.A.
(2003). Mercury effects of predator avoidance
behavior of a forage fish, golden shiner (Notemigonus Crysoleucas).
Environmental Toxicology and Chemistry, 22, 7, 1556-61.
Weis, J.S. & Weis, P. (1998). Effects of exposure to lead on behavior of
mummichog (Fundulus heteroclitus L.) larvae .Journal of Experimental
Marine Biology and Ecology, 222, 1, 1, 1-10.
Williams F.E., White D. & Messer Jr, W.S. (2002). A simple spatial alternation task
for assessing memory function in zebrafish. Behavioural Processes.
58,125–132
Willner, P. (ed.). (1991). Behavioural Models in Psychopathology: Theoretical,
Industrial and Clinical Perspectives. Cambridge: Cambridge University
Press.
Wolach, A.H., McHale, M.A., Fitzpatrick, C.S. & Dunlap. R. (1975) Survival time
and
weight
loss
in
goldfish
(Carassius
auratus).
Psychological
Reporter,36, 3, 979-85.
Xavier, G.F. (1993). A modularidade da Memória e o sistema nervoso Central.
Psicologia USP, 4, 1/2, 61-116.
Zangrossi Jr., H. (1996). Modelos animais de ansiedade. Em F.G. Graeff & H.
Zangrossi Jr. (org). Ansiedade e Transtornos de Ansiedade. Rio de
Janeiro: Editora Cientifica Nacional.
70
Zella, W. & Wolach, A.H. (1980). Reversal and nonreversal shifts in a goldfish
(Carassius auratus) classical conditioning situations. The Psichological
Record, 30, 25-37.
71
ANEXO I – Mini-laboratório para peixes
72
Materiais:
•
2 lâmpadas de aquário, completas (com calha, reator, starter);
•
5 metros de cortina Black out.
•
4,5 metros de trilho pra cortinas e acessórios (rolinhos, gancho,
derlimitadores)
•
1 timer
•
2 tomadas de extensão de 4 plugs
•
5 metros de fio para tomadas
•
1 plug macho para tomada
•
4 aquecedores
•
um areador de 4 saidas ou 4 filtros externos de 60 l
•
4 aquários de vidro de 35x20x25 cm
•
aquários de teste conforme descritos anteriormente no texto
•
1 aquário de vidro de 30 x 30 x 20 cm (para uso como campo aberto)
•
Rack feito em madeira resistente ou metalão 25;
o Para ser feito de madeira
4 caibros de madeira de 5 x 5 x 10 cm
8 caibros de madeira de 5 x 5 x 60 cm.
parafusos para fixar e cola
3 madeiras (pode ser compensando revestido) de 2,5 x 55 x
55 cm
73
1 madeira compensada revestida de 130 x 55 x 2,5 cm
4 roldanas para geladeira resistentes.
O desenho esquemático segue nas próximas paginas.
74
55 cm
2,5 cm
1
60 cm
Frontal
2.5 cm
2
130 cm
60 cm
2,5 cm
3
75
2,5 cm
1
60 cm
2.5 cm
2
130 cm
60 cm
2,5 cm
3
Lateral
76
2,5 cm
60 cm
Traseira
2.5 cm
130 cm
60 cm
2,5 cm
77
55 cm
peça 1
superior
55 cm
55 cm
inferior
55 cm
78
55 cm
peça 2
superior
55 cm
55 cm
inferior
55 cm
79
55 cm
peça 3
superior
55 cm
55 cm
inferior
55 cm
5cm
colunas
pés
10 cm
4x
8x
60 cm
80
eletrica
plug
vista traseira
Timer
p/lampada 1
p/ tomada 1
p/lampada 2
P/ tomada 2

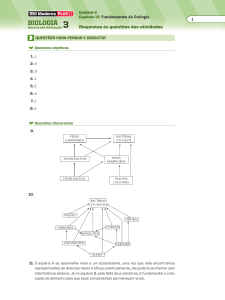







![[#0365] Construção de aquário suspenso](http://s1.studylibpt.com/store/data/001003998_1-a54b836971d985ae5004abb9aca75a09-300x300.png)


